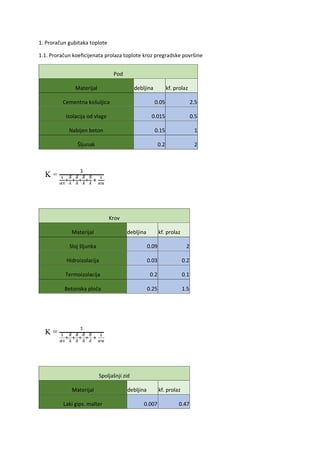
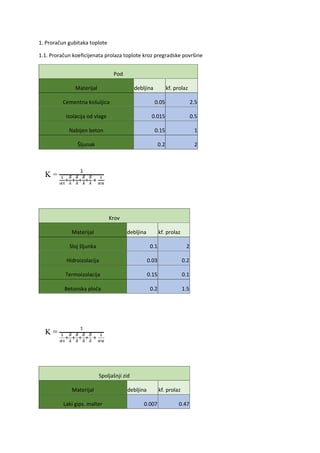
Genetika bakterija mliječne kiseline 261
C. Bakterijski hromosomi
Geni koji šifriraju sve esencijalne, katabolitičke i biosintetičke aktivnosti ćelija
se nalaze u hromosomima. Zato znanje o strukturi i organizaciji hromosoma u
bakterijama mliječne kiseline ima fundamentalan značaj i primjenljivu
vrijednost za mliječnu industriju. Nedavna napredovanja u tehnologiji
mapiranja hromosoma i u izvorima sekvenciranja nukleotida je potaknulo
interest za analizu genoma bakterija i hromosomi mliječnih bakterija sigurno
nisu iznimka.
Napore da se karakteriziraju hromosomi bakterija mliječne kiseline su ranih
1970-ih i 1980-ih poduzeli naučnici koji su koristili kinetiku renaturacije DNK-
DNK da procijene veličinu genoma (u daltonima) na
En. Faecalis, Lc.lactis
i
patogenim streptokokoma (Bak i suradnici, 1970; Jarvis i Jarvis, 1981). Klasične
metode za izmjenu gena kao što je transdukcija i konjugacija (vidjeti dio III.A i
III.C) nisu prikladne za mapiranje hromosoma kod bakterija mliječne kiseline,
tako da detaljnije studije genoma nisu bile izvodljive do pronalaska tehnologije
pulsirajućege električno polja gel-elektroforezom (PFGE) ranih 1980-ih godina
(Le Bourgeois i suradnici, 1993). Ova metodologija omogućava da se pročiste
relativno nedirnuti bakterijski hromosomi, digestiraju sa rijetkim rezanim
ograničenim endonukleazama, a onda riješi problem proizvoda ograničenja
velike molekularne težine elektroforezom u obrnutom električnom polju. Ako
se koriste standardi odgovarajuće veličine u gelu, sumiranje individualnih
restriktivnih fragmenata nakon PFGE postaje brzo i relativno tačno sredstvo za
procjenu veličine genoma. Ovim pristupom prikupljene su procjene veličina za
sojeve više od 15 vrsta bakterija mliječne kiseline. Ovi podaci pokazuju da
bakterije mliječne kiseline kao druge nutritivno zahtjevne eubakterije imaju
relativno male (približno 1.8 do 3.4 megabazna para) hromosome (Davidson i
suradnici, 1996). Jedna od praktičnih opažanja nastala ovim radom jeste da su
polimorfizmi restriktivnih fragmenata zajednički za PFGE profile iz različitih
sojeva istih vrsta bakterija mliječne kiseline. Ovaj pronalazak je doveo do toga
da se PFGE koristi kao oruđe za uzimanje otisaka DNA za identifikaciju sojeva i
za evaluaciju roda sojeva (Le Borugois i suradnici, 1993). Drugi važan ishod
PFGE tehnologije je bila njegova upotreba, kombiniranjem sa drugim
procedurama kao što je Južna hibridizacija sa specifičnim genskim sondama,
da se prikupe skromne fizičke i genetičke mape hromosoma bakterija mliječne
kiseline.
Ova strategija je korištena da se naprave mape za hromosome nekoliko
industrijski važnih bakterija mliječne kiseline, uključujući
Lc.lactis
(Davidson i
suradnici, 1995; Le Borugois i suradnici, 1992a; Tulloch i sur., 1991),
O.oeni
(Ze-
Ze i sur., 1998) i
S.thermophilus
(Rousell i sur., 1994), i za mnoge pategnske
streptokoke (Dimitrij i sur., 1998; Gasc i sur, 1991; Hantman i sur, 1993;
Suvorov i Fereti, 1996).
Ove mape su potvrdile da se kod individualnih vrsta pa čak i sojeva mogu
razlikovati organizacija i veličina genoma, i pokazuju da sve bakterije mliječne
kiseline karakterizirane do danas imaju jedinstven i cirkularan hromosom.
Konačno, PFGE tehnologija je također pomogla u proučavanju geometrije
hromosoma i intraspecifičnih polimorfizma kod
Lc.lactis
i
S.thermophilus.
Ova
istraživanja su identificirala intraspecifične genomske polimorfizme koje su
nastale inverzijom, umetanjem, brisanje i translokacijom DNK i pružila su dokaz
da su IS elementi bili uključeni u mnoge od ovih radnji (Davidson i sur., 1996;
Leblond i Decaris, 1998; Roussel i sur., 1997). Kako je primijećeno u dijelu II.B
dodatna istraživanja su potvrdila da su velike inverzije genoma kod hromosoma
Lc.lactis
ML3 ustvari nastale homolognom rekombinacijom između IS
905
elemenata.
Kako je naznačeno u prethodnim paragrafima razvoj i komercijalizacija PFGE
tehnologije je pomogla nastajanje nove mikrobiološke discipline čiji je predmet
interesa strukturalna, funkcionalna i komparativna analiza bakterijskih
genoma.
Iako je PFGE analiza još važan dio istraživanja genoma najuzbudljiviji i
inovativniji rad u ovom brzo rastućem polju sada potiču analize nukletoidskih
sekvenci potpunih genoma.
1. Komparativna genomika
Kompiliranje i označavanje cjelokupne sekvence genoma je revolucioniziralo
bakteriologiju i mikrobialnu genetiku i stvorilo je skoro nezamislive prilike da
se istraži bakterijska evolucija, genetika, psihologija i metabolizam. Od 1995
godine je objavljen cjelokupni nukleotidni niz za više od 30 različitih
mikrobialnih genoma i izvode se projekti sekvencioniranja za preko 100
drugih vrsta ( vidjeti http:/ /www.tigr.org/tdb/mdb/mdb.html). Dramatičan rast
u istraživanju sekvence genoma je uveliko rezultat tehnoloških poboljšanja
kod automatiziranih DNK sekvencera, alata molekularne biologije, ličnih

D. Bakteriofagi
Bakteriofagi, ili skraćeno fagi, su virusi koji napadaju i uništavaju bakterijske
ćelije. Inhibitorni efekat koji ovi obligatorni paraziti imaju na bakterije
mliječnih startera je poznat više od 60 godina, i njihov destruktivni utjecaj na
industriju sira i jogurta je fokusirao pažnju širom svijeta na molekularnu
genetiku i evoluciju faga bakterija mliječne kiseline. Pošto industrijska
fermentacija sa starterima
Lc. lactis
i
S. thermophilus
trpi velike ekonomske
gubitke trenutno razumijevanje biologije faga bakterija mliječne kiseline
uveliko potječe od faga koje inficiraju ove dvije vrste (Brussow i sur., 1998;
Garvey i sur., 1995). Međutim nekoliko grupa je opisalo da bakteriofagi
inficiraju druge industrijski važne vrste bakterija mliječne kiseline uključujući
mnoge mliječne laktobacile, i neki od ovih faga su čak karakterizirani do nivoa
genomskog sekvenciranja (Alterman i sur., 1999; Koadira i sur., 1997;
Mikkonen i sur., 1996). Takonomijski, nekoliko faga sa uvlačivim repovima-
bičevima (familija Myoviridae) ili veoma kratkim repovima (familija
Podoviridae) je izolirano iz bakterija mliječnih kiselina, ali većina bakteriofaga
koje inficiraju ove vrste pripadaju familiji Siphoviridae (fagi sa neuvlačivim
repovima) iz reda Caudovirales (Brüssow i sur., 1998; Caldwell i sur., 1999;
Davis i sur., 1985; Dı´az i sur., 1992;
Garcı´a i sur., 1997; Jarvis i sur., 1991, 1993; Manchester, 1997; Park i sur, 1998;
Se´chaudi sur., 1988; Trevors i sur., 1983).
Detaljan opis morfologije faga bakterija mliječne kiseline, infektivnih krugova
svojstava raspona domaćina je predstavljen u Poglavlju 6 ove knjige i o njima
se ovdje neće više govoriti. Umjesto toga, ovaj dio će naglasiti neke od
postojećih rezultata molekularno genetskog istraživanja bakteriofaga bakterija
mliječne kiseline.
Za razliku od hromosoma bakterija mliječne kiseline, gdje obećanje genomskog
istraživaja ostaje nepotvrđeno, evolucionarna studija na genomima
bakteriofaga bakterije mliječne kiseline je ubrzano napredovala zadnjih
nekoliko godina. Očigledan razlog za ovu razliku je taj da su genomi faga
mnogo manji (veličina varira od 18 do 134 kb) (Prevots i sur., 1990), od
bakterijalnoh hromosoma, i zbog toga mogu biti puno brže sekvencionirani (i
puno jeftinije).
Dva najvažnija rezultata studija genetike i genoma faga uključuju (1) iscrpan
pogled na različitost i evoluciju bakteriofaga bakterija mliječne kiseline i (2)
aplikacija elemenata iz faga da se poboljša otpor bakteriofaga kod mliječnih
bakterija startnih kultura i za genetsku manipulaciju ovih vrsta.
1. O porijeklu faga
Dizajn učinkovitih strategija za kontrolu faga za mliječnu fermetativnu
industriju u velikoj mjeri ovisi o poznavanju diverziteta i evolucije
bakteriofaga. Zbog toga je porijeklo faga u tvornicama mlijeka bilo predmet
velikog istraživanja i debata, i jedna od glavnih tačaka ove diskusije je bila
uloga lizogenije u evoluciji virulentnih faga. Kako je naglašeno prije (vidjeti dio
II.B.1), Shimizu-Kadota i kolege (1985) su pokazali da avirulentna faga
Lb.casei
jasno potiče od profaga u starter bakterijumu domaćina sa insertivnim
prenošenjem ISL
1
. Otkriće da je lizogenija poprilično uobičajena kod mliječnih
bakterija mliječne kiseline, a naročito kod
Lc.lactis
je dovela do spekulacije da
profagi mogu biti važan rezervoar litičkih bakteriofaga mliječne industrije
(Davidson i sur., 1990). Sada znam da iako virulentni
Lc. lactis
mogu da nastaju
od umjerenih faga (Davidson i sur., 1990), većina litičkih i umjerenih faga koje
inficiraju ovu vrstu dijele veoma malo DNK homologije i zbog toga nisu blisko
povezani (Garvey i sur., 1995).
Važan izuzetak uključuje litičke fage vrste P335, koje pokazuju DNK
homologiju sa umjerenim bakteriofagima a u fabrikama sira im se učestalost
povećava (Dumaz i Klaenhammer, 2000; Moineau i sur., 1994; Walker i sur.,
1998).
Značajnije, novi P335 litički fagi evoluiraju uz pomoć hromosomske DNK
domaćina, i analiza sekvence nukleotida jednog od tih fagmenata je potvrdila
da potiče od komponenata profaga (Dumaz i Klaenhammer, 2000; Moineau i
sur., 1994).
Za razliku od situacije kod
Lc.lactis
, sve litičke i umjerene
S.thermophilus
bakteriofage karakterizirane do danas pripadaju jednoj DNK homolognoj grupi
(Brussow i sur., 19989 i komparativna genomikaje ustanovila da brisanja kod
lizogenskog modula umjerenih faga vjerovatno igraju ključnu ulogu u evoluciji
litičkih faga (Lucchini i sur., 1999a; Tremblay i Moineau, 1999). Nasreću, čini se
da je lizogenija jako rijetka kod ovih vrsta (Le Marrec i sur., 1997).
Lizogeni su češći kod mliječnih laktobacila (Davidson i sur., 1990), međutim,
ustanovljena je i genetska veza između litičkih i umjerenih faga od nekih od
ovih vrsta (Auad i sur., 1999; Lahbib-Mansais i sur., 1988; Mikkonen i sur.,
1996; Shimizu-Kadota sur., 1985). U cjelosti, ovi podaci pokazuju da lizogenija
ima važnu (ali ne ekskluzivnu ) ulogu u evoluciji novih litičkih faga u industriji
mliječne fermentacije, i kako tvrde za razvoj startnih bakterija mliječne
kiseline tretiranih profagima(Shimizu-Kadota i Sakurai, 1982).
Gledajući pobliže, studije komparativne genomike bakterija mliječne kiseline
Siphoviridae su također dale dobar uvid u taksonomiju i evoluciju
bakteriofaga. Kao što je tipično za fage sa repovima, sve genomi faga bakterija
mliječne kiseline opisani dosad imaju linearne, dvostruke DNK molekule čiji je
sadržaj G_C paralelan onom kod domaćina (Ackerman 1999). Ovisno od
mehanizma kojima su smješteni u kapsidu (koji može da ovisi čak i između
blisko pvoezanih bakteriofaga) genomi bakterija mliječne kiseline Siphoviridae
posjeduju kohezivne krajeve ili kružnu permutaciju sa terminalnim zalihama.
Za većinu ORF-a faga se čini da su prepisane iz zajedničke grupe, osim kod
umjerenih faga, gdje se skup gena povezan sa lizogenijom divergentno

morfogenetskih modula lamboida i bakterija mliječne kiseline Saphoviridae ne
može biti izvučena da se dodijele zamišljene funkcije mnogim genima faga
bakterija mliječne kiseline (Chandry i sur.,1997; Desiere i sur., 1999).
Nedavna potvrda ove strategije od strane Desierea i suradnika (1999) bi trebala
ohrabriti istraživanje strukture i funkcije kod genoma faga bakterija mliječne
kiseline koje će na kraju pružiti uzbudljiv uvid u biologiju bakteriofaga
bakterija mliječne kiseline i interakcija između faga i domaćina.
2.
Novi alati za biotehnologiju bakterija mliječne kiseline
Istraživanje genoma bakteriofaga je također doprinijelo nastajanju novih
mehanizama za odbranu od faga za mliječne startne kulture. Prvi sistem koji
će biti opisan uključuje umetanje replikacije bakteriofagskog porijekla u
prevozni vektor streptokoka (Hill i sur., 1990). Laktokoke ćelija domaćina koje
su nosioci rekombinantnih plazmida posjeduju abortivni fenotip otporan na
fage nazvan Per ( za otpornost uslovljenu fagima) za kojeg se vjeruje da djeluje
titracijom replikacijskih proteina faga od izvornih
ori
sekvenci faga tokom
ranih stadija infekcije (Hill i sur., 1990; McGrath i sur., 1999). Iako je otpornost
na fage preko Per originalno ustanovljena kod
Lc.lactis
nedavna istraživanja
Foleya i njegovih suradnika (1998) sugerišu da Per sistemi možda zapravo
imaju veću vrijednost kod
S.thermophilus.
Razlozi za ovo su dupli: prvo, veoma
je malo prirodnih sistema otpora na fage dostupno za ove vrste; i drugo, čini se
da Per sistemi imaju relatinvo široku otpornost na fage
S.thermophilusa
(Foley
i sur., 1998). Drugi primjeri sistema otpora na fage dobivenih iz genetike
bakteriofaga uključuju (1) aplikaciju protismislene mRNA protiv vrlo očuvane
sekvence faga
Lc. Lactis
(Kim i Batt, 1991; Walker i Klaenhammer, 2000); (2)
sistem za
Lc.lactis
koji stavlja samoubilački gen pod kontrolu strogo fagi-
inducibilnih promotera da bi se usmrtile ćelije domaćina tokom infekcije
( Đorđević i sur., 1997); i (3) mehanizam koji daje imunitet superinfekciji
umjerenih faga kod
Lb.casei
konsitutivnom ekspresijom domaćina na gen faga
za protein represor (Alvarez i sur., 1999).
Funkcionalna genomska analiza faga bakterija mliječne kiseline je također
donijela niz korisnih alata za molekularno-genetsku manipulaciju startera
bakterija kod mlijeka (Venema i sur., 1999). Naprimjer, integraze gen (
int
) i
pričvšćenje sekvence/niza (
attP
) koji posreduju integracijom umjerenih faga
određenog mjesta u hromosom domaćina su osposobljeni da razviju vektore
integracije koji unose stranu DNK na određenu lokaciju (
attB
) na
bakterijalnom hromosomu.
Int-attP
integracijski sistemi pružaju nekoliko
važnih prednosti preko ekvivalenata koji se oslanjaju na homolognu
rekombinaciju u posredovanju domaćina.
To uključuje (1) integracija se dešava na
attB
, lokacija koja se inače koristi za
unos profaga i zbog toga je manje moguće da omete ćelijske funkcije ili život;
(2) stabilnost integranta je pod neselektivnim uvjetima obično visoka; i (3)
očuvanje
attB
sekvence kod drugih bakterija, ili fleksibilnost u njenom