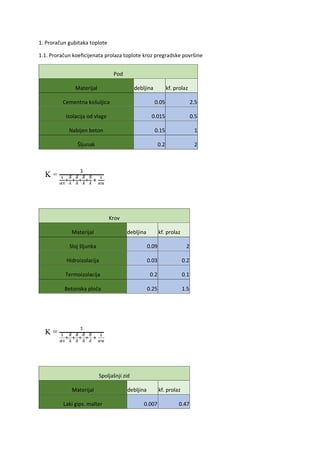
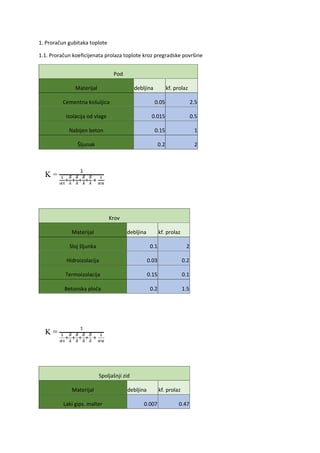
ODSEK EKOLOGIJA
Seminarski rad iz ''EKOLOGIJA BILJAKA''
Matrice u populacijama višegodišnjih zeljastih
perena
Milica Rakić
378072 / 2012
Datum: 01. februar, 2014.
Poena: ______
Dr. Jasminka Đ., Miloradović, prof.
____________________________
Požarevac
Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
2014.
2

Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
Sadržaj
1. Uvod ......................................................................................................................– 4 –
1.1. Lesli-jev matrični model ........................................................................ – 5 –
1.2. Leftković-ev matrični model ..................................................................– 6 –
2. Perene ....................................................................................................................– 8 –
3. Matrice kod višegodišnjih perena ......................................................................... – 8 –
3.1. Matrice u populacijama monokarpnih perena (biena) ........................... – 8 –
3.2. Matrice u populacijama polikarpnih perena .......................................... – 11 –
4. Zaključak ...............................................................................................................– 16 –
5. Literatura ...............................................................................................................– 17 –
4
Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
1. Uvod
Iako je matematika već dugo vremena isprepletena s biološkim istraživanjima,
eksplozivna sinergija između matematike i biologije javila se tek u novije vreme.
Najvažniji razlozi tome su:
intenzivan razvoj računala koji omogućavaju izvođenje komplikovanih
matematičkih računa i simulacija, što u prošlosti nije bilo moguće,
povećan interes za modeliranje, ponajviše zbog etičkih problema, rizika i
nepouzdanosti, te drugih komplikacija povezanih s istraživanjem ljudi i drugih
živih bića, i
nedavan razvoj novih matematičkih metoda, kao npr. teorije kaosa koja pomaže u
razumevanju kompleksnih, nelinearnih mehanizama u biologiji.
Otkriće mikroskopa u drugoj polovini 17. veka uzrokovalo je revoluciju u
biologiji. Otkriće mikroorganizama zahvaljujući izumu mikroskopa, dovelo je do naglog
razvitka biologije, te nastajanja novih bioloških grana. Matematika se u određenom
smislu može smatrati „novim mikroskopom“ koji može otkriti, inače nevidljive svetove u
raznim vrstama podataka.
Današnji biolozi sve više uviđaju da je uz pomoć prikladne matematike moguće
interpretirati razne vrste podataka. U tom smislu matematika je novi biološki mikroskop,
samo bolji (Cohen, 2004).
Biološke nauke već dugo vremena su inspiracija za razvoj novih matematičkih
metoda, koje pomažu u objašnjavanju kompleksnih bioloških fenomena. Biologija će
stimulirati nastanak fundamentalno nove matematike, budući da je živi svet kvalitativno
puno heterogeniji nego neživa priroda. U tom smislu, biologija je nova fizika
matematike, samo bolja (Cohen, 2004).
Primena matematike u biologiji imala je znatan uticaj na razvoj novih grana
matematike, kao i na razvoj novih metoda koje su potrebne za obradu bioloških podataka.
To se na prvi pogled može činiti neobičnim, prvenstveno zbog različite prirode
matematike i biologije. Matematika inzistira na strogosti i preciznosti, matematičke
činjenice su nepromenjive, a uspešne matematičke teorije imaju životni vek od nekoliko
hiljada godina.
5

Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
Leslie-eva projekciona matrica L definiše na koji način se jedinke u populaciji
kreću kroz starosne grupe. U prvom redu projekcione matrice L nalaze se vrednosti
fertiliteta (
Fi)
pojedinih starosnih grupa (
i
), odnosno prosečan broj potomaka po ženki u
određenom vremenskom intervalu. Na poddijagonali se nalaze verovatnoće
preživljavanja (
Pi
), odnosno verovatnost da će jedinke starosne grupe
i
preći u skupinu
i+1
. Na ostalim mestima u matrici nalaze se vrednosti 0 (2):
(2)
Množenjem projekcione matrice s populacionim vektorom koji predstavlja
sadašnji broj jedinki u svakoj dobnoj skupini, dobijamo vrednost veličine populacije u
sledećem vremenskom intervalu (Caswell, 2001). Iterativnim postupkom, dobijamo
veličinu i strukturu populacije nakon nekog izabranog vremenskog intervala.
Vrednosti potrebne za izračunavanje parametara projekcione matrice
izračunavamo iz tablica preživljavanja i tablica uzrasne strukture (eng.
life tables
). Kako
te vrednosti često nisu dostupne, parametre je potrebno proceniti na druge načine.
1.2. Leftković-ev matrični model
Modifikacija Leslie-evog matričnog modela koja populaciju opisuje godinama,
predložio je 1965 Lefkovitch. U tom modelu, jedinke su podeljene u stadijume, koji
mogu biti različitog trajanja. U određenom projekcijskom intervalu jedinke mogu preći u
sledeći stadijum, ili ostati u istom stadijumu. Lefkovitch-eva matrica stoga ima oblik (3):
(3)
7
Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
Vrednosti u prvom redu matrice (
Fi
), označavaju fertilitet pojedinih stadijuma,
odnosno prosječan broj potomaka po ženki u projekcijskom vremenskom intervalu.
Vrednosti na poddijagonali (
Pi
), predstavljaju verovatnoću da će jedinka koja se nalazi u
stadijumu
i
, u sledećem projekcionom intervalu preći u sledeći stadijum,
i+1
. Vrednosti
na glavnoj dijagonali matrice (
Gi
), predstavljaju verovatnosti da će jedinka određenog
stadijuma
i,
u promatranom projekcijskom intervalu, ostati u istom stadiju.
Prilikom konstruiranja ovog modela, bitno je naglasiti da je trajanje stadijuma
nezavisno od vremenskog projekcionog intervala, u kojem se mere parametri matrice.
Bez obzira na trajanje projekcionog intervala i trajanje stadijuma, parametri matrice se
izračunavaju kao broj događaja po projekcionom intervalu (Caswell, 2001).
Kao slikoviti prikaz projekcijskih matrica koriste se grafikoni životnog ciklusa
(eng.
life cycle graphs
). Stadijumi ili starosna doba prikazuju se kružićima (čvorovima).
Verovatnoće prelaska u sledeći stadijum ili starosnu grupu označavaju se
vodoravnim strelicama. Verovatnoća opstanka u stadijumu tokom projekcionog intervala
prikazuju se kružnim strelicama. Fertiliteti pojedinih stadijuma ili dobnih skupina
označeni su polukružnim strelicama koje vode natrag do prvog stadijuma ili prve starosne
grupe.
Sl. 1. Slikoviti prikaz Lefkovitch-eve projekcione matrice (eng.
life cycle graph
). Jedinke tokom
projekcionog intervala mogu ostati u stadiju (
Gi
) ili preći u sledeći stadijum (
Pi
). Razmnožavaju se samo
odrasle jedinke (
F
).
8

Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
narušenih i otvorenih staništa zahvaljujući
rezervi semena, kao što su deponije
najrazličitijih materija iz kamenoloma ili
rudnika uglja i minerala.
Životni ciklus biena i konačna stopa
populacionog rasta se mogu uspešno generisati
Leftković-evim matricama, jer je u mnogo
slučajeva dokazano da dužina života i prelazak
iz vegetativne u reproduktivnu fazu zavise od
veličine rozete ili količine asimilata
translociranih u korenu i zavise od opštih
uslova rastenja i razvića. Za populaciju
Verbascum phlomoides
koja kolonizuje
biološki prazan prostor (deponije pepela uglja)
Djordjević-Miloradović & Miloradović (2000)
su pokazali da se opstanak i širenje populacije zasniva na produkciji semena i klijaju više
puta tokom godine. U populaciji je nađeno osam uzastopnih klasa: semena, klijanci, četiri
razvojna stadijuma virginilnih pre-reproduktivnih rozeta i dva stadijuma cvetalih, veće i
manje. Konačna stopa populacionog rasta je veća od jedinice (λ = 1.101). Nadalje je
perturbaciona analiza pokazala koji element matrice najviše utiče na konačnu stopu
populacionog rasta.
Ova analiza je pokazala da na lamda najčiše utiče verovatnoća da semena klijaju,
dok su ostali elementi manje značajni. Elastični obrazac ove matrice pokazuje nadalje
najveći udeo upravo navedenog matričnog elementa, ali ni ostale verovatnoće nisu bez
značaja. Produženo trajenje životnog ciklusa istražene biena koja cveta u drugoj, trećoj,
četvrtoj ili čak petoj godini života, usled nepoviljnih uslova za rastenje na pepelu ima za
posledicu funkcionalnu zavisnost izmešu veličine rozete vegetativnih rozeta i broja
produkovanih semena kod generativnih adulta (Djordjević-Miloradović & Miloradović,
2002.)
Lofgren et al (2000) istraživali su populacionu dinamiku
Carlina vulgaris
u
kasnom sukcesivnom stanju na jalovini. Na osnovu 8 projekcionih matrica koje su
Sl. 2 -
Verbascum phlomoides
10
Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
istražili za eksperimentalne i
prirodne populacije našli su da je u
samo dve populacije λ>1, pri čemu
elastičnost pokazuje najveći značaj
verovatnoće da semena klijaju ili
elementa
a
21
. Na opstanak i širenje
populacija
Carlina vulgaris
ključni
uticaj ima povremeno narušavanje
staništa i otvaranje bioloških praznih
prostora. Sa razvojem sukcesivne
zajednice
Carlina vulgaris
pokazuje smanjenje u brojnosti do isčezavanja.
Na osnovu rezultata ispitivanja
populacije
Dipsacus sylvestris
Caswell
(1989, 2001) je uradio brojne
simulacije matrične analize i ispitivanja
konačne stope populacionog rasta,
elastičnost i sensitivnost. U populaciji
Dipsacus sylvestris
opisano je sedam
uzastopnih klasa: semena (s),
dormantna semena stara 1 godinu,
dormantna semena stara dve godine,
male rozaste biljke, srednje rozete,
velike rozete i cvetale biljke. Na osnovu tradicionalnih verovatnoća predložena je matrica
koja sadrži i verovatnoće klijanja aktuelnih semena, dormantnih semena starih 1 i 2
godine:
Sl. 3. Carlina vulgaris
Sl. 4.
Dipsacus sylvestris
11

Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
U populacijama jagorčevine (
Primula vulgaris
)
u šumama pri različitom svetlosnom rezimu opisano je
pet uzrasnih stanja (klijanci – Sd, juvenili – j, mali –
A
1
, srednji – A
2
i veliki adulti – A
3
.
Matrična analiza pokazuje da konačna stopa
populacionog rasta varira od vrednosti 1-2 u jednom
periodu istraživanja do 0.8-1.3 u drugom periodu.
Znači, populacije pokazuju varijabilnost u vremenu.
Senzitivna analiza pokazuje da je cvetanje i
reprodukcija najvećih adulta najosetljiviji element u
životnom ciklusu populacija, a da se osetljivost ovog
stanja povećava sa zatvaranjem krošnji šumskog
drveća i pogoršavanjem svetlosnog režima staništa.
Elastičnost pokazuje najveće vrednosti u delu životnog ciklusa koji se odnosi na
fekunditet.
Tokom šest godina (1982-1987) Horvitz & Schemske (1995) su proučavali
populacije
Calathea ovandensis
u četiri različite sredine definisane u odnosu na tip
podloge, svetlosni režim i gustinu biljaka. Na osnovu ovakvih demografskih parametara
konstruisana je matrica iz koje se vidi da je
istražena vrsta sa dugovremenom
semenskom bankom, veoma varira u broju
pojave klijanaca (10 puta se razlikuje broj
klijanaca iz godine u godinu), ima veoma
nizak mortalitet reproduktivnih adulta, a
fekunditet se povećava sa veličinom biljaka.
Konačna stopa populacionog rasta varira od
0.73 do 1.25. Za period od sešt godina
što ukazuje na povoljne i stabilne
uslove u kojima istražene populacije
C.
ovandensis
živi. Nadalje, populaciona stopa
rasta je u korelaciji sa semenskom
Sl. 6. Sanicula europeae
Sl. 7.
Calathea ovandensis
13
Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
produkcijom, klijanjem semena i rastom klijanaca (na osnovu elastičnosti). Selekcija u
ovim populacijama deluje na stanje klijanaca i juvenila stanja.
Kod nekih vrsta perena pojava, dužina života i populacioni rast zavise od
periodičnog narušavanja i destrukcije staništa kao što su pojava požara. Istražujući pojavu
i opstanak nekih retkih, ugroženih perena u prerijama SAD-a posle požara, kao što je
Lomathium bradshawii
, Koye (2001) je primenio matričnu analizu. U životnom ciklusu
Lomathium bradshawii
je našao semena, klijance, vegetativne biljke sa 1, 2, 3 listova i
reproduktivne biljke sa 1, 2, 3 i više cvasti u obliku štita.
Upoređujući matrice dobijene na osnovu posmatranja promena u populacijama na
različitim mestima u preriji Oregona (SAD) koja u prošlosti nisu pretrpele požar ili je
požara bilo u bližoj ili daljom
prošlosti, Koye nalazi da
populacije na požariptu imaju
λ>1 ili λ=1, dok populacije bez
narušavanja staništa požarom u
poslednjih šest godina imaju
λ<1.
Nadalje je elastična
analiza pokazala da u smanjenu
populacionog rasta (kada je
λ<1) najvećeg udela ima tranzicija da vegetativne sa tri lista ne odrastaju u cvetale kao i
da mlade reproduktivne ne rastu i ne cvetaju, već da duže ostaju u tzv. Stanju statisa. U
slučaju povećanja u populacionoj veličini (posle 2-3 tretmana požarom) najveću
elastičnost pokazuju elementi reprodukcine koji su označeni kao tranzicija R1 u R2 i R2
u R3. Zaključak je da je za opstanak i širenje populacija
Lomathium bradshawii
neophodno povremeno narušavanje prerije požarom, koji se i inače periodično javlja u
prerijama, jer sa smanjenjem biološki praznog prostora smanjuje se i uslov za rast
Lomathium bradshawii
, a raste rizik isčezavanja zbog brze pojave superiornijih
kompetitorskih vrsta biljaka.
U demografiji šumske invazivne perene
Prosopis glandulosa
korišćena je tzv.
Jednogodišnja i periodična matrica. U životnom ciklusu je uočena pojava da su pojedine
Sl. 8.
Lomathium bradshawii
14

Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
Konačna stopa populacionog
rasta populacije
T. farfara
koja
kolinizuje pepeo je veća od jedinice i
iznosi λ = 1.321. Elastična analiza i
sumarno izražavanje elastičnosti po
stanju ili klasi pokazuje da najvećeg
značaja u populacionom rastu ima
pojava juvenilnih na dva načina, iz
klijanaca i vegetativno, kao i pojava
mladih virginilnig izdanaka iz rizoma.
Formiranje gap-a ili otpvaranjem praznog prostora u šumama stimulativno deluje
na grananje i vegetativno razmnožavanje mahovine
Hylocomium splendens
. Prema
matričnom modelu za trogodišnji period istraživanja λ = 1.012 za kontrolu populacija, a
posle tretmana prosvetljavanja u eksperimentalnim populacijama λ = 1.740, dok isecanje
grana ili segmenata mahovina utiče na opšte smanjenje populacije (λ = 0.841). Grafik
životnog ciklusa koji korespondira projekcionim matricama predstavlja rast mahovine u
kontrolnoj i tretiranim populacijama. Populaciono povećanje kod prosvetljenih, ali ne
narušenih populacija se zasniva na sposobnosti zrelih ogranaka mahovine da prežive
ovakav tretman i grananjem daju nove jedinke. Ovakva analiza pruža odgovor na bitno
pitanje populacionog ponašanja mahovina u narušenim šumskim sistemima i njihovoj
sposobnosti za regeneraciju i obnavljanje.
U populacionoj dinamici perene
Achnatherum calamagrostis
na deponijama
jalovine uočeno je da se busenovi jedinki razvijaju kao klijanci, juvenilni ili odrasli u
zavisnosti od broja izdanaka koje sadrže. Konačna stopa populacionog rasta izračunata je
na osnovu broja kvadrata koje okupiraju busenovi
Achnatherum calamagrostis
tokom
dve godine. Na taj način uočeno je da u životnom ciklusu ove perene postoji stanje
unutrašnjeg razvoja ili dinamika razvoja genetsa i kolonizacije vegetativnim rastenjem. U
svim istraživanim populacijama λ>1. Veće povećanje pokazuju populacije koje se
razmnožavaju i iz semena i vegetativno, nego one koje imaju samo vegetativni rast.
Sl. 10. Tussilago farfara
16
Ekologija biljaka
– Matrice u populacijma višegodišnjih zeljastih perena
4. Zaključak
Perene ili trajnice predstavljaju zeljaste, ređe zimzelene baštenske biljne vrste sa
životnim vekom od 2-20 godina. Areal rasprostranjenja je cela planeta, odnosno svaka
klimatska zona obiluje visegodisnjim cvetnicama karakteristicnim za dati region. Mnoge
vrste su samonikle, pa cak i korovi, ali vecinu predstavljaju kultivisani ukrasni varijeteti
osnovne vrste. Odlikuje ih brz rast i razvoj, skromni zahtevi prema uslovima sredine i
zemljišta, dugotrajnost i visoka dekorativnost.
U odnosu na habitus, odnosno oblik stabla delimo ih na četiri osnovne kategorije:
- perene sa jastučastm habitusom,
- perene sa končastim habitusom,
- perene sa žbunastim habitusom,
- perene sa puzavim habitusom.
Pored toga, kod nekih vrsta se javljaju odstupanja u pravilnosti cvetanja, najčešće
uslovljena klimatskim (vremenskim) uslovima. Ponekad se dešava ponovljeno cvetanje,
kad vrsta cveta u aprilu-maju, pa ponovo u oktobru, dok se kod nekih vrsta cvetanje
produžava i traje od aprila do oktobra.
Razmnožavanje perena može biti generativno i vegetativno.
Generativno razmnožavanje podrazumeva setvu semena, kad se dobijaju mlade
biljke koje cvetaju tek druge-treće godine (nekad i kasnije).
Velika raznolikost prema veličini, obliku, boji cveta i u odnosu na uslove koje
zahtevaju, brz razvoj i visoka dekorativnost čine ovu grupu biljaka obaveznim elementom
pri formiranju zelene površine. Bilo da se perene koriste kao pokrivači tla, za vertikalno
ozelenjavanje, ili kao centralni detalji u cvetnjaku (perenjaku), u kombinaciji sa
kamenom, vodom ili drugim biljkama, perene su nezamenljive na zelenoj površini.
Dobro poznavanje pojave, preživljavanja, uzrasne strukture i drugih karaktera
populacija perena je danas neophodno. Ako biljke u populacijama prolaze kroz određena
onogenetska stanja onda je stanje razvijenosti značajnije od uzrasta ili starosti.
17