1
SINTEZA PROTEINA
Proteini predstavljaju složene makromolekule nastale me usobnim spajanjem aminokiselina u
polipeptidni lanac po ta no utvr enom rasporedu. Informacija o na inu njihove sinteze sadržana je u
genima, odre enim odsje cima DNK molekule. Veza izme u DNK i sinteze proteina objašnjena je kroz
procese
replikacije, transkripcije
i
translacije
tzv. centralnom dogmom molekularne biologije.
Najjednostavnije re eno,
osnova navedene dogme je da se informacije sadržane u DNK preslikavaju
posredstvom RNK u strukturu proteina.
Da bi se razumjela sinteza proteina neophodno je poznavati gra u DNK molekule i op enito nukleinskih
kiselina, te gra u ribosoma.
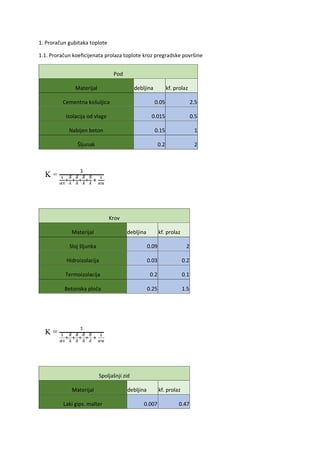
Gra a DNK molekule
Molekula
DNK
izgra ena
je
iz
dva
polinukleotidna lanca koji su me usobno povezani
vodikovim vezama i uvijeni oko zajedni ke
vertikalne osi. Kraj polinukleotidnog lanca na
kome je slobodna fosfatna grupa na 5-C atomu
ozna ava se kao 5' kraj, a suprotni kraj je ozna en
kao 3'. Unutar dvostruke zavojnice navedeni lanci
su antiparalelni, odnosno pružaju se u suprotnim
smjerovima, što zna i da se nasuprot 5' kraja
jednog lanca uvijek nalazi 3' kraj drugog lanca i
obrnuto.
Svaki polinukleotidni lanac zasebno predstavlja linearni polimer ija je osnovna gra evna jedinica
nukleotid. Strukturu svakog nukleotida ine jedna azotna baza, jedna molekula še era i jedna molekula
fosforne kiseline. Postoje dva tipa azotnih baza koje mogu biti sastavne komponente nukleotida: purinske
(adenin, gvanin) i pirimidinske baze (citozin, timin i uracil). U sastavu DNK nukleotida mogu se nalaziti
adenin, gvanin, citozin i timin, dok se kod RNK nukleotida umjesto timina javlja uracil. Razlike izme u
ovih nukleotida o itavaju se i u še ernoj komponenti. Še er koji ulazi u sastav DNK nukleotida je
deoksiriboza, dok je sastavni dio RNK nukleotida riboza.
Nukleotidi su unutar polinukleotidnog lanca me usobno povezani fosfodiesterskim vezama koji povezuju
C
3
- atom jednog nukleotida i C
5
- atom susjednog nukleotida. Ovi dijelovi polinukleotidnog lanca,
fosfatne skupine i še erna komponenta, ine okosnicu svake molekule nukleinske kiseline. Hidrofilni su,
te su stoga orijentirani prema izloženoj, vanjskoj strani zavojnice. Naspram tome, azotne baze koje ovim
slijedom vezivanja ostaju slobodne, zbog hidrofobnih interakcija orijentiraju se prema unutrašnjosti
zavojnice. Ovakvim postavljanjem molekula u prostoru vidljivo je da su baze jednog polinukleotidnog
lanca uvijek okrenute prema bazama nasuprotnog polinukleotidnog lanca. Povezivanje dva lanca ostvaruje
se uspostavljanjem vodikovih veza izme u purinske baze jednog lanca i pirimidinske baze drugog lanca.
Pri tome se adenin uvijek veže sa timinom, a gvanin sa citozinom, ukoliko je rije o DNK molekuli.
Samim tim o ito je da redoslijed baza u jednom lancu u potpunosti odre uje redoslijed baza u drugom
komplementarnom lancu. Ovo pravilo, koje se ozna ava kao komplementarnost baza, od velikog je
zna aja prilikom elijske diobe i prenošenja nasljednog materijala jer omogu uje identi no prenošenje
nasljednog materijala iz elije majke u elije k eri.
2
Gra a RNK molekule
Molekula RNK je duga ka, jednolan ana molekula koja služi kao posrednik u
prenošenju genetske informacije sa DNK molekule do ribosoma gdje se u
kona nici odvija sinteza proteina. U hemijskom pogledu gra a RNK
molekule je vrlo sli na gra i DNK molekule. Sastoji se od RNK nukleotida
me usobno povezanih u polinukleotidni lanac, a svaki RNK nukleotid u
svom sastavu sadrži: purinske baze adenin i gvanin, pirimidinske baze citozin
i uracil, še er ribozu i fosfornu kiselinu. U usporedbi sa DNK nukleotidom,
azotna baza timin zamijenjena je uracilom, a še erna komponenta
deoksiriboza ribozom.
S obzirom na ulogu u sintezi proteina razlikuju se tri vrste RNK molekula:
Informacijska RNK
(iRNK)
- prenosi genetsku informaciju sadržanu u
DNK molekuli iz elijskog jedra do ribosoma. Sastavljena je od dugih nizova
ribonukleotida. Budu i da redoslijed nukleotidnih tripleta u iRNK kodira i
redoslijed aminokiselina u polipeptidnom lancu jasno je da e se pojedine
iRNK me usobno znatno razlikovati s obzirom na dužinu lanca i molekulsku
masu. Ovisno o vrsti proteina za iju sintezu su odgovorni, molekulske mase
im se kre u od sto tisu a do nekoliko milijuna.
Ribosomska RNK
(rRNK)
- izgra uje ribosom i njegova je osnovna sastavna komponenta uz proteine.
Molekulske mase rRNK su velike i sadrže veliki broj nukleotida.
Transportna RNK
(tRNK)
- prenose aminokiseline do iRNK na ribosomima, niske su molekulske mase i
sastoje se od svega 75 do 90 nukleotida.
Ribosomi
Ribosomi su hidratizirane, nemembranske organele ve inom kuglastog ili sferi nog oblika, promjera 15 -
30 nm. Njihova osnovna funkcija u eliji usko je vezana uz sintezu proteina.
Približno 60% hemijskog sastava ribosoma ini ribosomska RNK, dok ostatak predstavljaju proteini. Broj
ribosoma u eliji nasljedno je odre en dok je njihov raspored ovisan o vrsti elije, fazi njenog rasta i
razvi a. Najve im dijelom su smješteni u citoplazmi, gdje mogu biti slobodni ili vezani za endoplazmatski
retikulum. Nalaze se i u jedru, plastidima i mitohondrijima.
Ribosomi ne napuštaju svoju životnu sredinu, iako bi s obzirom na svoju veli inu mogli pro i kroz pore
membrana nekih od navedenih organela. Veli ina ribosoma izražava se koeficijentom sedimentacije ili
Svedbergovom konstantom koja predstavlja mjeru za veli inu molekula dobivenoj na osnovu brzine
taloženja pri centrifugiranju. Brzina taloženja ne ovisi samo o masi estice, ve i o njenom obliku.
Vrijednosti brzine sedimentacije za ribosome iznose od 1 - 200 x 10¹³s. Zbog jedostavnijeg prikaza
rezultata navedene vrijednosti se iskazuju u Svedbergovim jedinicama (S) pri emu jedna Svedbergova
jedinica iznosi 1 x 10¹³s.
Iako je princip njihove gra e sli an, ribosomi se u prokariotskoj i eukaritskoj eliji znatno razlikuju.
Prokariotski ribosomi su lakši, njihov sedimetacijski koeficijent iznosi 70 S, a razbacani su po cijelom
protoplastu. Sedimentacijski koeficijent eukariotskih ribosoma je nešto ve i i iznosi 80 S. U eukariotskoj
eliji ribosomi se ne nalaze samo u citosolu ve i u matriksu mitohondrija i plastida, a najve im dijelom
vezani su za membrane endoplazmatskog retikuluma koji uslijed toga poprimaju hrapav izgled. Ribosomi
smješteni u organelima svojim izgledom i veli inom više podsje aju na prokariotske ribosome.