SKELETNI MIŠIĆI
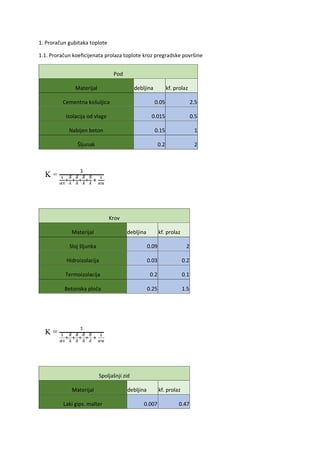
Skeletni mišići su efektori somatskog nervnog sistena. Oni izvršavaju naredbe koje putem motornih
neurona stižu iz različitih delova CNS.
Svojstva mišićnih ćelija
Mišićne ćelije imaju osobinu
kontraktilnosti
zahvaljujućl prisustvu kontraktilnog aparata. Druga
osobina im je
ekscitabilnost
, tj membrana mišićne ćelije može da generiše i sprovodi akcioni potencijal.
Raspored jona u mišićnoj celiji i izvan nje sličan je kao kod neurona, pa su i električna zbivanja i jonski tokovi u
mišiću u osnovi isti kao kod nervne ćelije. Razlike postoje u intenzitetu i trajanju promena. Potencijal
mirovanja membrane mišećne ćelije je oko
-9 0 mV
.
Mišići u celini imaju i osobinu
elastičnosti.
Elastičnost omogućava mišiću da se posle kontrakcije
vrati na dužinu u mirovanju. Ova osobina je naročito važna kod skeletnih mišića uključenih u disanje.
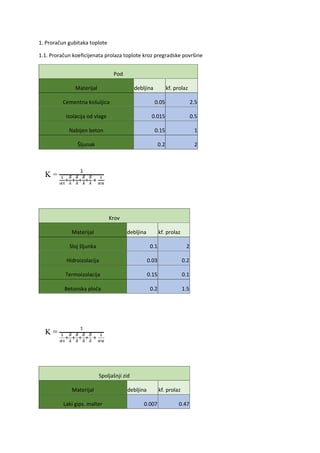
Nervno mišićna sinapsa
U fiziološkim uslovima skeletni mišić se kontrahuje samo ako je stimulisan motornim neuronom. To
je eferentni neuron koji potiče iz sive mase CNS, prednih rogova sive mase kičmene moždine za skeletne mišiće
trupa i ekstremiteta ili iz motornih jedara glavenih živaca za skeletne mišiće glave i vrata. Motorni neuroni su Aα
tipa.
Veza između motornog neurona i mišićne ćelije je nervno-raišićna sinapsa. Transmiter je Ach i ova
sinapsa je uvek ekscitatorna.
Membrana mišićnog vlakna ispod aksonskog završetka je modif ikovana i naziva se
završna
(motorna)
ploča.
Ona je udubljena i u tom udubljenu koje se zove
primarna sinaptička
pukotina
leži
aksonski završetak. Pošto je aksonski završetak veoma mali, a mišićno vlakno može da bude vrlo dugačko,
površina sinaptičkog kontakta se povećava formiranjem invaginacija sarkoleme u primarnoj sinaptičkoj
pukotini i nastaje
sekundarna sinaptička pukotina
.
Ach oslobođen iz presinaptičkog završetka motornog neurona difunduje kroz sinaptičku pukotinu
do receptora na motornoj ploči a to su receptori nikotinskog tipa. Vezivanjem dva molekula Ach za α
subjedinice receptora otvara se kanal koji propušta Na, K, ali i Ca jone. Neto efekat je umerena depolarizacija
regiona završne ploče. Ta depolarizacija se naziva
potencijal završne ploče
i iznosi na oko -15 mV.
Potencijal završne ploče je prolazan, jer delovanje ACh kratko traje. Ach razlaže acetilholinesteraza
pričvršćena za bazalnu laminu sarkoleme u sekundarnoj sinaptičkoj pukotini.
Membrana završne ploče ne može da generiše akcioni potencijal zato što ne sadrži voltažno-zavisne
kanale za Na odgovorne za generisanje akcionog potencijala. Potencijal završne ploče se zato širi pasivno do
susednih segmenata sarkoleme koji ne učestvuju u sinapsi i kada se njihov membranski potencijal smanji do
nivoa praga nastaje akcioni potencijal. Amplituda akcionog potencijala skeletne mišićne ćelije je slična
amplitudi akcionog potencijala neurvnog vlakna, samo duže traje. Akcioni potencijal se propagira slično kao
kod nemijeliniziranih nervnih vlakana. Pošto se nervno-mišićna sinapsa nalazi na sredini mišićne ćelije, akcioni
potencijal se sarkolemom širi na obe strane.
Akcioni potencijal tokom svog prostiranja aktivira kontraktilni mehanizam i taj proces se
naziva:
IX i X predavanje 2006. B. PLEĆAŠ
1
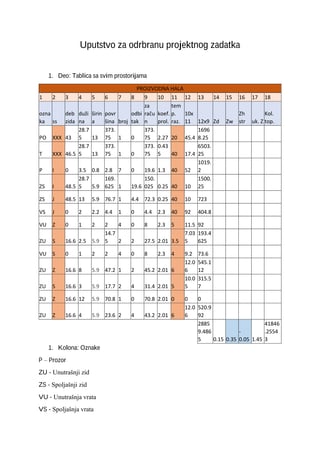
Povezivanje ekscitacije i kontrakcije
Proces kojim depolarizacija membrane mišićnog vlakna dovodi do kontrakcije naziva se
povezivanje ekscitacije i kontrakcije.
Akcioni potencijal koji se širi sarkolemom, prelazi i na sistem T-tubula,
jer one predstavljaju invaginacije sarkoleme, otvara kanale za Ca u njihovoj membrani i Ca iz ECT u
neposrednoj blizini ulazi u ćeliju. Međutim, ključna promena nije taj ulazak Ca, već promena konformacije
kanala za Ca u membrani T-tubula. Promena konformacije tih kanala izaiva promenu u konformaciji drugog
tipa kanala za Ca u membrani sarkoplazminog retikuluma u neposrednoj blizini i oslobađanja velikih količina
Ca iz sarkoplazminog retikuluma, tako da se intracelularna koncentracija ovog jona se povećava oko 1000x.
Povećanjem koncentracije Ca započinje kontrakcija, a njen intenzitet e srazmeran koncentraciji Ca u
citosolu. Ca se odmah vezuje za troponin C, što ima za posledicu pomeranje tropomiozina sa aktina i otkrivanje
vezujućih mesta na aktinu za glave miozina. Tada mogu da se uspostave
poprečni mostovi
ili
veze
između
miozinskih glava i subjedinica aktina. Veza se uspostavlja prvo pod uglom od 90° u odnosu na aktin. Formiranjem
poprečnih mostova istovremeno se aktivira ATP-aza u miozinskim glavama i hidrolizuje ATP do ADP i PO4 .
Energija koja se pri tome oslobađa omogućava obrtanje miozinske glave i smanjenje ugla poprečne veze na 45°,
što opet rezultuje klizanjem tankih aktinskih filamenata po miozinu. Pošto su filamenti aktina jednim krajem
pričžvršćeni za Z linije koje predstavljaju granice sarkomere, sarkomere se skraćuju, a time i mišićno vlakno.
Poprečni most se zatim cepa i ako ima dovoljno ATP i Ca, miozinska glava uspostavlja novu vezu sa sledećim G-
aktinom. Jedno uspostavljanje poprečne veze, smanjivanje ugla i cepanje veze naziva se
ciklus poprečnih
veza.
Jedan ciklus skraćuje mišić svega oko 1%. Aktivno skraćivanje mišića i do 60% izazvano je brzim
uspostavljanjem i cepanjem velikog broja sukcesivnih veza.
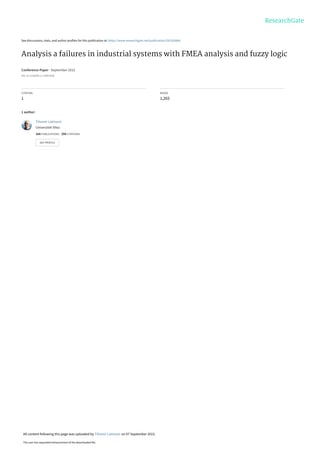
Odmah nakon oslobađanja Ca iz sarkoplazminog retikuluma počinje suprotan proces - vraćanje Ca
u cisterne sarkoplazmatičnog retikuluma. Povećanje koncentracije Ca u sarkoplazmi aktivira pumpu Ca-ATP-
azu koja ubacuje Ca u sarkoplazmin retikulum i, ukoliko ne stigne sledeći akcioni potencijal, vrlo brzo se
uspostavlja relaksirano stanje. U sarkoplazminom retikulumu nalaze se Ca-vezujući proteini
kalsekvestrin
i
kalretikulin
koji vezuju veliki broj jona Ca i na taj način smanjuje koncentraciju slobodnog Ca u unutrašnjosti
sarkoplazminog retikuluma, pa i gradijent koncentracije nasuprot koga pumpa u membrani sarkoplazminog
retikuluma ubacuje Ca. Kada se koncentracija Ca u sarkoplazmi smanji ispod jedne kritične vrednosti, prekida
se veza između aktina i miozina, relaksirajući proteini se vezuju za aktin i nastupa relaksacija. Energija je
neophodna i za relaksacju, odnosno za rad pumpe koja transportuje Ca u sarkoplazmin retikulum. Jedan
molekul ATP se troši za transport 2 jona Ca. Ako energije za ovaj proces nema, ne dolazi do relaksacije mišića.
Ekstremni primer je
rigor mortis
-
mrtvačka ukočenost.
Karakteristike mišićne kontrakcije
Iako je uobičajeno da se termin kontrakcija povezuje sa skraćivanjem mišića, mišić može da se
kontrahuje i bez promene dužine.
Postoje 2 vrste mišićne kontrakcije: izotonusna i izometrijska kontrakcija.
Izitonusna kontrakcija
je ona tokom koje se mišić aktivno skraćuje. Takva kontrakcija se javlja za
vreme hodanja, trčanja ili podizanja.
Izometrijska kontrakcija
je ona kod koje se mišić kontrahuje nasuprot sile opterećenja, bez
smanjenja dužine, ali uz povećanje napetosti (tenzije). Javlja se kada opterećenje ili sila koja se opire skraćivanju
izjednači sa snagom mišićne kontrakcije.
Prosta i složena mišićna kontrakcija
IX i X predavanje 2006. B. PLEĆAŠ
2