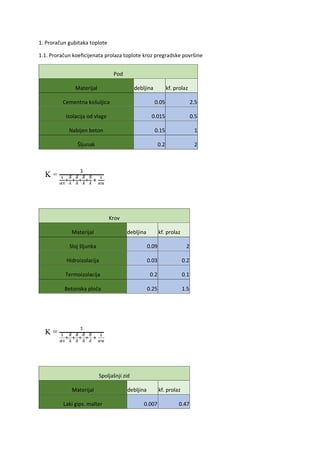
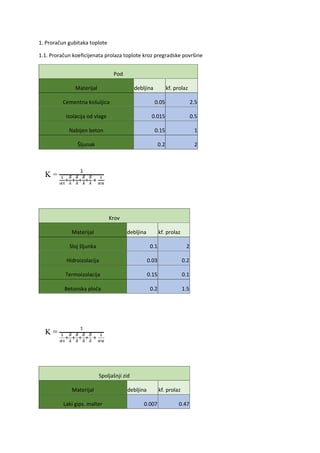
SKRIPTA
ZAŠTITA DRVETA OD KSILOFAGNIH INSEKATA
(za studente odseka za šumarstvo)
Ova skripta sadrži delove knjige Zaštita drveta I deo (ksilofagni insekti) autora prof. dr Konstantina
Vačića (1971) i namenjena je za spremanje ispita iz predmeta Zaštita drveta koga polažu studenti odseka za
šumarstvo koji su predhodno položili ispit iz predmeta Šumarska entomologija.
2

svojim hodnicima. Kod larava drvotočaca kretanje potpomažu i čekinje čiji su vrhovi okrenuti
unazad.
Kod larava nekih ksilofagnih insekata javljaju se posebni organi koji služe za zbijanje
crvotočine ili za njeno izbacivanje iz hodnika. Tako na primer kod lađara (Lymexylidae) i kod osa
drvenarica (Siricidae) postoje na kraju tela jaki izraštaji pomoću kojih one obavljaju
navedene
radnje. Nije isključeno da ovi dodaci u izvesnoj meri služe i za lakše kretanje u hodnicima.
Dalje pojave koje su vezane sa prilagođavaljem načinu života su rudimentacija čulnih organa i
depigmentacija kože. Od prvih naročito su redukovane oči koje u tami hodnika ne vrše nikakvu
funkciju. Slepe larve ksilofagnih insekata ipak razlikuju svetnost i tamu (najvetovatnije preko kože),
jer njihovi hodnici nikada ne izlaze na površinu napadnutog drveta.
Depigmentacija kože je takođe posledica života u tami. Koža ksilofagnih insekatskih larava
najčešće je bezbojna, ili ima žućkastu, odnosno prljavo belu nijansu. Ređe se javljaju crvenožute ili
izrazito voštano žute forme. To je slučaj kod insekaita čije larve u toku razvoja više puta menjaju
sredinu u kojoj se razvijaju (na primer gusenice drvesnice - Zeuzera pyrina L. prelaze iz grančica u
grane, da bi na koncu završni deo svoga razvića obavile u stablu; gusenice vrbotočca Cossus cossus L.
često napuštaju stabla u kojima su se izlegle i odlaze bilo u druga stabla, bilo u zemlju radi čaurenja).
Pored opisanih spoljašnjih, javljaju se i izvesne adaptacione karakteristike unutrašnjih delova
tela. Od ovih da na prvom mestu pominjemo građu prednjeg želuca (proventriculus). Kod većine
ksilofagnih formi on je jako razvijen, snabdeven vrlo snažnom muskulaturom, a njegova unutrašnja
hitinsika opna obrazuje zadebljanja u vidu rebara, zubaca i sl. Ogledima je ustanovljeno da ovaj
organ virši pulzirajuće pokrete i da doprinosi finijem usitnjavanju hrane, odnosno drveta (navedeni
autor unosio je u proventrikulus parafin, a zatim ga je vadio putem disekcije i našao na njemu otiske
zubaca i rebara). Da proventrikulus služi finijem usitnjavanju hrane dokazuje i to što su čestice drveta
u srednjem crevu sitnije nego u proventrikulusu i što kod insekata koji nemaju razvijen
proventrikulus (strižibuba - Xystrocera globosa) čestice drveta u srednjem crevu imaju znatno veće
dimenzije nego u slučajevima gde je prednji želudac razvijen (na primer kod Macrotoma palmata
drvo je u obliku finog praha).
Pored ovoga javljaju se specifičnosti u građi drugih delova crevnog aparata. Ovde na prvom
mestu treba pomenuti rektalnu kesu termita - proširenje zadnjeg creva u kome su smeštene
simbiontske prazivotinje (bičari), dalje fermentacionu komoru larava listorožaca (Lamellicornia) u
kojoj žive bakterije čiji fermenti razlažu celulozu. Prošireno zadnje crevo ovih insekata ima na
unutrašnjoj strani delove na kojima je kutikula snabdevena jako razgranatim čekinjama; kutikula
između ovih je jako tanka i prožeta je finim kanalićima. U ovim regionima zadržavaju se čestice
drveta vrlo dugo, sve dok ih mikroorganizmi ne razlože. Verovatno je da se ovde vrši i apsorpcaja
hrane. Organi koji sadrže simbiontske mikroorganizme (micetomi), nalaze se i na zidovima srednjeg
creva mnogih drvotočaca (Anobiidae), i strižibuba (Cerambycidae). Osim toga simbionti mogu da se
nalaze i u posebnim ćelijama srednjeg creva. Micetomi se dalje nalaze i u masnom telu i sl.
Adaptivne osobine unutrašnjih organa ksilofagnih insekata još su vrlo malo proučene i
nesumnjivo je da će nova istraživanja otkriti niz drugih koje su rezultat života u drvetu i ishrane
drvnom masom.
FIZIOLOGIJA ISHRANE I VARENJA KOD KSILOFAGNIH INSEKATA
U poglavlju o hemijskom sastavu drveta videli smo da se ono sastoji od različitih materija i da
je biološka vrednost tih supstanci različita. Glavnu masu drveta čine celuloza i lignin, dakle jedinjenja
koja su hemijski vrlo stabilna i koja životinjski organizmi teško koriste (celuloza čini 60—62% suve
supstance drveta a lignin 18—38%). Drvene polioze dolaze na drugo mesto po zastupljenosti
(pentozani 6—23% i heksozani 2—14%). U daleko manjoj meri susreću se lakše svarljive materije
(skrob 0—5%, šećeri 0—6,2% i belančevine 1,1—2,3%). Iz izloženog pregleda se vidi da je drvo kao
hranjiva supstanca vrlo bogato u teško svarljivim ugljenim hidratima, ali ujedno vrlo siromašno u
lako svarljivim ugljenim hidratima i belančevinama. Stoga je od naročitog interesa rešiti pitanje na
koje načine insekti koriste teško svarljive ugljene hidrate i kako se snabdevaju supstancama u
minimumu (belančevine, vitamini i sl.). U ovom poglavlju ćemo glavnu pažnju obratiti na
4
iskorišćavanje i varenje celuloze. Lignin koji se takođe nalazi u znatnim količinama u drvnoj masi
insekti uopšte ne mogu da koriste. Da je ovo tačno svedoči i ogled u kome su termiti hranjeni drvetom
čija je prethodna hemijska analiza ustanovila da sadrži 54,6% celuloze, 18% pentozana i 27,4%
lignina. Izmet ovih insekata sadržavao je svega 18% celuloze, 8,5% pentozana i čak 75,5% lignina. Iz
navedenih podataka se vidi da je najveći deo pentozana i celuloze iskorišćen, dok je lignin kao potpuno
nesvarljiv koncentrisan u izmetu i izbačen napolje.
VARENJE CELULOZE
Već smo pomenuli da ima vrlo malo životinja koje koriste celulozu direktno. Naprotiv,
fermenat celulaza daleko je rasprostranjeniji kod mikroorganizama, i to pretežno onih koji pripadaju
biljkama (bakterije i gljivice). Sve do najnovijeg vremena verovalo se da insekti koriste celulozu samo
posrednim putem, tj. pomoću simbiontskih mikroorganizama. Međutim, noviji radovi opovrgli su ovo
mišljenje i dokazali da insekti predstavljaju i u ovom pogledu izuzetak, tj, da su u stanju da vare
celulozu i sopstvenim fermentima. Ipak, kod znatnog broja vrsta nije nađena celulaza, a sposobnost da
vare celuloyu vezana je ustvari za prisustvo simbionata. Ovo je eksperimentom za termite nesumnjivo
dokazano. Prema tome postoje dva načina korišćenja i varenja celuloze: direktno i indirektno.
Direktno varenje celuloze. Mali je broj ksilofagnih insekata za koje je do danas uzvrđeno da su
sposobni da sami razlažu celulozu do glukoze, tj. da proizvode fermente celulazu i celobiazu. Pomenuti
fermenti nađeni
su u crevnom soku larava velike hrastove strižibube (Cerambyx cerdo L.), kućne
strižibube (Hylotrupes bajulus L.), Macrotoma palmata, riđe kućne strižibube (Stromatiium fuilvum)
šarenog drvtočca (Xestobium rufovillosum), tačkastog drvotočca (Anobium punctatum De Geer) i dr.
Naročito je aktivna celulaza larava riđe kućne strižibube. Pri PH = 5,5, prema Mansour-u njihov
crevni sok razlaže celulozu čak i onda kada je vezana sa ligninom, dakle u njenom najotpornijem
obliku.
Da navedeni insekti zaista vare celulozu svedoči i činjenica da njihov izmet sadrži daleko
manje celuloze i pentozana nego drvo u kome žive, odnosno i kojim se hrane. Za šarenog drvotočca
eksperimentom je dokazano da asimilira oko jednu trećinu ukupne celuloze koju pojede zajedno sa
ostalim sastojcima drveta. Lignin naprotiv uvek ostaje u istoj količini, te se koncentriše u izmetu.
Međutim, postoje i insekti koji uopšte nisu sposobni da koriste celuloze. Takav je slučaj sa
larvama buba beljikara (Lyctidae), kapucina (Bostrychidae), nekih strižibuba (na pr. Xystrocera) i
gusenicama vrbotočca (Cossus cossus). Glavnu hranu ovih insekata čine ekstraktivne materije, pre
svega skrob i šećeri, a u izvesnoj meri i drvne polioze (gusenice vrbotočca proizvode jedan fermenat
koji razlaže heksozane). Pošto se ove supstance nalaze u drvetu u malim količinama, razumljivo je da
larve navedenih insekata moraju da pojedu nesrazmerno velike količine hranljivog supstrata da bi
podmirile svoje potrebe u njima. Kod vrsta iz familije Lyctidae glavnu ulogu igraju ekstraktivne
materije koje se mogu direktno asimilirati (glukoza). Ako drvo ne sadrži šećere nego samo skrob, one
mogu dugo da žive, ali više ne rastu. Čim se hrani dodaju šećeri, porast se nastavlja a razviće znatno
ubrzava. Slična pojava konstatovana je i kod gusenica vrbotočca. Ako se hrane drvetom njihovo
razviće se završava tek u trećoj godiini. Međutim, hranjene šargarepom, završavaju razviće za svega
godinu dana. U laboratoriji Zavoda za entomologiju uspelo se dobiti leptire već osam meseci nakon
polaganja jaja. Verovatno je da razviće ubrzava ne samo veća količina šećera, nego i druge hranljive
materije, pre svega belančevine.
Indirektno korišćenje celuloze zastupljeno je kod velikog broja ksilofagnih
insekata. Najlepši
primer za ovo nalazimo upravo kod termita. Da su ovi insekti nesposobni da sami vare celulozu,
dokazano je eksperimentom. Ako se termiti drže 24 časa na tempeiraturi od 36°C ili pri povišenom
pritisku kiseonika (3,5 atm.), odnosno ako duže vreme gladuju, njihova crevna flora propada, te više
ne mogu da žive na čistoj celulozi, niti pak mogu da koriste celulozu drveta. Ovako sterilizovani insekti
mogu da žive još najviše 2-3 nedelje, a zatim propadaju od gladi iako im je crevni kanal pun hrane.
Međutim, ako je celuloza prožeta micelijama gljivica oni se ponašaju normalno i ostaju u životu. N a
koncu, ako im se obnovi crevna mikroflora (davanjem partikula izmeta drugih termita), insekti
ponovo postaju sposobni da žive na čistoj celulozi, odnosno da koriste celulozu drveta. Slični ogledi, sa
5

Belančevine pripadaju grupi hranjivih materija koje su neophodne za normalnu ishranu svih
živih bića nesposobnih da ih sintetišu iz neorganskih sastojaka. Prema tome i ksilofagni inseikti
moraju za svoj život i razviće imati na raspoloženju dovoljne količine ovih supstanci. Pošto je drvna
masa, kao što smo već videli, jako siromašna belančevinama (naročito srčika) njihove larve
zadovoljavaju potrebe svoga organizma u proteinskim materijama bilo tako što uzimaju nesrazmerno
velike količine drveta, bilo opet putem simbioze sa makroorganizmima.
U prvom slučaju u pitanju su najčešće insekti čije se larve razvijaju u beljici jer ovaj sloj
sadrži najviše belančevina. Konstatovano je međutim da ni svi delovi beljike nemaju jednake količine
ovih supstanci. Njima su najbogatiji slojevi u neposrednoj blizini kore. Ovo utiče na brzinu razvića
raznih vrsta ksilofaga na
taj način što one koje žive u plitkim slojevima beljike, imaju po pravilu
znatno kraći životni ciklus. Koliko količina belančevina utiče na brzinu razvića najbolje su pokazali
ogledi koje je izveo Becker. Navedeni autor je konstatovao da dodatak peptona drvetu u kome se
nalaze larve kućne strižibube mnogostruko ubrzava njihovo razviće. Isto tako je ustanovio da se larve
brže razvijaju u beljici, dok srčiku, siromašnu beiančevinama izbegavaju. Slične pojave konstatovane
su kod drvotočaca, ali samo u početnim stupnjevima larvenog razvića, jer kasnije oni svoje potrebe u
belančevinama zadovoljavaju pomoću simbionata (larve kućne strižibube nemaju simbionte).
Snabdevanje belančevinaima putem simbioze veoma je rasprostranjeno kod ksilofagnih
insekata. Videli smo da postoje vrste koje vare same simbionte (termiti), pošto ovi iskoriste celulozu i
druge sastojke drveta i sintetizuju od njih materije svoga organizma, koje su pristupačne insektima.
Ova pojava nije ograničena samo na endosimbionte. Na sličan način snabdevaju se belančevinama
insekti koji gaje ambrozija gljive u svojim hodnicima (Scolytidae, Platypodidae, delom Lymexylidae
itd.).
Vrste ksilofagnih insekata koje se snabdevaju belančevinama iz drveta u velikom stepenu su
zavisne od količine ovih materija u hranjivom supstratu. Tako na primer larve kućne strižibube koje
kao što smo videli nemaju simbionte, ne mogu da žive dugo na čistoj celulozi. Naprotiv, termiti i
Anobiidae imaju ovu sposobnost. Stoga su ove životinje u stanju da napadnu vrlo staro drvo u kome
sigurno više nema ni traga belančevina.
Poseban slučaj predstavljaju insekti truloga drveta. Njihove larve unose u sebe razloženu
drvnu masu, ali ne koriste njene sastojke, nego se ishranjuju isključivo micelijama gljivica koje bogato
prožimaju drvo u razlaganju. Stoga je i opravdan naziv koji Chauvin daje ovoj grupi insekata (lažne
ksilofage).
Samo varenje belančevina vrši se u srednjem crevu. Pod uticajem naročitih fermenata koji se
nazivaju proteaze belančevine se najpre raspadaju na albumoze i peptone, ovi na polipeptide koji pod
uticajem drugih encima (polipeptidaze) daju aminokiseline. Od raznih aminokiselina, pošto budu
apsorbovane u krv i transportovane do tkiva, insekti građe sebi specifične belančevine.
SNABDEVANJE VITAMINIMA
Eksperimentalno je ustanovljeno da su za normalno razviće insekata potrebni vitaimini B
grupe (B1, B2, B6, pantoteinska kiselina i dr.). Nedostatak navedenih supstanci izaziva zastoje u
porastu i druge poremećaje. Interesantno je da se ove pojave ne ograničavaju samo na životinje koje
su patile od avitaminoze, nego i na njihovo potomstvo, Drugi vitamini (A, C, D itd.) prema
dosadašnjim posmatranjima nemaju nikakvog značaja.
Poznato je da je drvo siromašno vitaminima, naročito ako se nalazi u vazdušno suvom stanju.
Stoga se i postavlja pitanje na koji način se ksilofagni insekti snabdevaju ovim neophodnim
supstancama.
Izvori vitamina su opet simbiontski mikroorganizmi, koji mogu pripadati najrazličitijim
grupama, a mogu se nalaziti u raznim organima. Kod drvotočaca i strižibuba nalaze se simbionti
(najverovatnije iz grupe kvasaca) u naročitim ćelijama srednjeg creva, odakle stalno prelaze u samu
šupljinu creva. Kakav je mehanizam korišćenja ovih mikroorganizama još nije u potpunosti poznato.
Najverovatnije je da insekti razlažu samo mrtve simbionte. Kod termita smo videli da koristeći bičare
7