Seminarski rad iz biomehanike
Alexander
(Animal Mechanics,
1968), i neurobiology Roberts
(Neurophysiology of
Postural Mechanisms,
1978). U ovog radu govorićemo o nervnim kontrolama, mehanici i
energetici pokreta. S jedne strane biće uglavnom reči o hodanju i trčanju na zemlji. S
druge strane, biće ograničen na one aspekte hodanja i trčanja koji su značajni za
razumevanje funkcionisanja lokomotornog sistema.
1.1. Hodanje: uporedne razlike
Ribe se kreću unapred tako što šalju talase ili savijaju unazad kičmeni stub. One ovo
čine periodičnim kontrakcijama mišića jedne strane tela dok su mišići druge stane nepomični.
Ovo je primenjeno na dati segmentni nivo: fazni pomak između rostralnog (prema glavi ) i
repnog segmenta (prema repu) obezbeđuje da putujući talas bočnog savijanja napreduje niz
telo. Ribe se kreću brže sa povećanjem učestalosti pomeranja repa. Brzina talasa nastalog
savijanjem raste linearno sa frekvencijom , tako da isti deo dužine talasa leži nasuprot telu ribe,
nezavisno od brzine talas
1
.
Kada su prsna i trbušna peraja postala prednji i zadnji udovi primitivnih
četvoronožaca, kretanje prvih kopnenih životinja se oslanjalo na pokrete riba. Svakako da
daždevnjak prikazan na slici 1. koristi bočno talasasto kretanje na zemlji koje veoma podseća
na kretanje koje koristi u vodi. U vodi daždevnjak pliva u osnovi slično kao i riba, sa
udovima pripijenim uz telo. Na zemlji, pokreti bočnog savijanja značajno produžavaju
dužinu koraka.
Iz nekih razloga, sisari ne talasaju bočno previše kada trče. Stoga što udovi kod sisara
nisu istureni sa strane kao kod primitivnijih vrsta, sisari ne bi puno toga dobili praveći
talasaste pokrete u transverzalnoj ravni. Umesto toga, oni savijaju i ispavljaju kičmu ali
samo tokom galopiranja kada se dva prednja uda pokreću skoro u fazi jedan s drugim kao i
što čine i zadnji udovi. Ova karakteristika sisara je iziskivala evolutivne promene u obliku
pršljenova. Način na koji su pršljenovi povezani kod vodozemaca i gmizavaca sprečava
uvrtanje kao i pokrete sabijanja i izduživanja kičmenog stuba. Kod sisara mnogi od ovih
dodataka koji nemaju sposobnost uvrtanja su izostali tako da dozvoljavaju ograničeno
uvrtanje i savijanje kičmenog stuba kako u bočnoj tako i u vertikalnoj ravni.
Kitovi i delfini savijaju i opružaju kičmu tokom plivanja kao što se pretpostavlja da su
i njihovi prastari preci to činili tokom trčanja na zemlji
2
.
1.2. Hodanje: klasifikacije
Hodanje dvonožaca.
Dvonošci imaju izbor da pokreću svoje donje udove bilo u fazi
(skakanje) ili u naizmeničnoj fazi (trčanje ili hodanje).
Torbari,uključujući kengure, skočimiše i miševe mogu da razviju prilično veliku
brzinu tokom skakanja. Postoji posebno interesantan odnos između brzine skakanja,
1
Grillner, 1975; Hertel, 1963
2
Grillner, 1975
4
Seminarski rad iz biomehanike
učestalosti koraka i potrošnje kiseonika kod kengura koji sugeriše da se oni kreću poput
rezonanznog harmonijskog oscilatora.
Hodanje karakterišu periodi tokom kojeg su obe noge na zemlji (dupli oslonac) nakon
kojih slede periodi stoja na jednoj nozi tokom koje se druga noga njiše unapred. Trčanje za
razliku od hodanja ne uključuje bilo kakav period dvostrukog oslonca. U stvari, tokom trčanja
postoji period često značajan kada su obe noge u vazduhu.
Hodanje četvoronožaca.
Broj različitih načina hodanja se kod četvoronožaca penje sa
3 na 6 (iako je često zgodno zanemariti neke razlike i govoriti samo o hodanju, kasu i
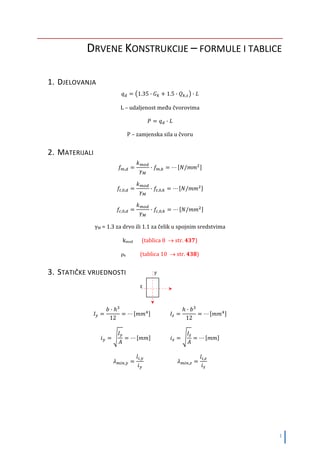
galopiranju). & načina kretanja su prikazani na tabeli 2 uz određivanje 2 parametra svake
noge (Mc Ghee,1968). Faktor se definiše kao deo ukupnog ciklusa koraka tokom kojeg je
noga na zemlji.
Neki autori više vole da odvoje naizmenično hodanje (na vrhu tabele 1) od faznog
hodanja (na dnu tabele 1). To predstavlja korisnu razliku zato što odvaja one korake koji ne
koriste značajnu fleksiju-ekstenziju kičme (naizmenično hodanje) od one vrste hodanja kod
kojih je to slučaj (fazno hodanje). Ta razlika će imati značajne posledice na zavisnost veličine
tela od učestalosti galopiranja.
Naizmenično hodanje se koristi pri malim brzinama; fazno hodanje pri velikim
brzinama.
5