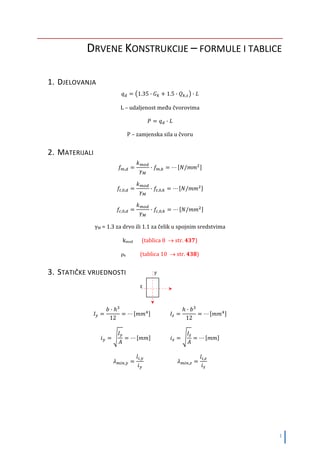
vlakna ne pokazuju osobinu poprečnog širenja prilikom uzdužnog skraćenja i obrnuto.
(Metsubara i Eliot, 1972), a sposobna su za aktivnu kontrakciju.
1.2. Elektrostatičke teorije
Logično objašnjenje uzroka klizećeg pokreta može biti sledeće – dva filimenta
prenose električni naboj suprotnog naelektrisanja, što čini da se međusobno privlače, pa se, s
toga, i površina njihovog preklapanja povećava. Ovaj mehanizam je prikazan u različitim
oblicima. (Ju i saradnici, 1970; Nobel i Polak, 1977).
E. F. Haksli ukazuje na nemogućnost ovakvih modela da se skrate više od one dužine
koja je potrebna za potpuno preklapanje filamenata aktina i miozina, ili na nemogućnost da
objasne smanjenje stope oslobođenja energije po jedinici promene dužine, kako se brzina
skraćivanja povećava. Takođe postoji i problem visoke koncentracije kalijum jona unutar
vlakna, koja prikriva naboje upućene negativno naelektrisanom filamentu.
1.3. Uvijanje tankih filamenata
Podolski (1959) je ukazao na mogućnost da se krajevi tankih filamenata najpre
pričvšćuju za susedne debele filamente, a zatim se tanki filamenti skraćuju, uvijajuće se u
zoni preklapanja. Ali, elektronskom mikroskopijom pokazano je da krajevi tankih filamenata
klize unutra prilikom kontrakcije, čak do tačke međusobnog preklapanja u centru „A“ trake.
Ovo je snažan dokaz protv teorije da se kontrakcija dešava zato što se sami filamenti
skraćuju.
1.4. Dokazi za postizanje nezavisnih generatora sile koji rade
ciklično
Ima dva posebno važna dokaza koji potvrđuju pretpostavku da su poprečne spojnice
izolovani generatori sile, koji deluju nazavisno.
1) Izometriski tetanički napon je proporcionalan rasponu (obimu) preklapanja aktina i
miziona. Gordanovi, Hakslijevi i Julijanovi elementi (1966) u kojima se koristio servo
kontrolisani uređaj za praćenje kako bi se održala određena dužina u ograničenom delu
preparata koji se sastoji od jednog vlakna, pokazuju da je izometrijski tetanički napon, koji
stvara vlakno, direktno proporcionalan rasponu preklapanja tankih i debelih filamenata, te je
s toga proporcionalan i broju aktivnih poprečnih veza, uzimajući u obzir jedinstveni poprečni
raspored.
Ovi podaci se takođe brojčano slažu sa trenutno najpoznatijim merama dužine debelih
i tankih filamenata. Ukupna dužina tankih filamena sa obe strane Z – linije u zgrčenim
vlaknima mišića žabe iznosi 1,98 μm (E.F.Hoksli, 1979).
1
Iako još uvek postoje određene
nedoumice u vezi sa dužinom debelih filamenata, trenutno najpouzdaniji dokazi koji su
1
Ova dužina filimenata u originalnim papirima (Gordn, A. F. Hazlej i Julijan, 1966), iznosila je 2,05 μm ali je
broj naknadno preinačen.
4
zasnovani na rengenskim difrakcionim ispitaivanjima, pokazuju da je dužina debelih
filamenata tokom izometrijske kontrakcije oko 1,67 μm (Haselgrouv, 1975). Uzimajući u
obzir ove vrednosti, procenjuje se da preklapanje aktina i miozina dostiže nultu vrenost kada
se mišić rastegne do dužine
capromere
1,98 + 1,67 = 3,65 μm. Ovo je vrlo blizu dužini na
kojoj je napon bio ravan nuli u eksperimetima iz 1966, kao što se očekivalo na osnovu
teorijske osnove.
2) Sledeća serija dokaza pokazuje da je brzina neoštećenog skraćenja nezavisna od
preklapanja aktina i miozina. Na slici 2. mišiću žabe omogućeno je da se skrati maksimalnom
brzinom prilikom opterećenja velikim teretom (manje od 3% tetaničkog napora). Kružeći na
ovoj slici pokazuju brzinu okraćivanja izmerenu sa opadajuće krive koja pokazuje odnos
brzina – vreme, na različitim rastojanjima. Kako je mišić počeo da radi kada je razmak
između pruga iznosio 3,1 μm, tačke na 3,0 μm i iznad ne treba uzimati u razmatranjnj jer je
mišić još uvek ubrzavao kada su one upisane (zabeležene). Zapanjujući zaključak je da
maksimalna brzina skraćivanja ne zavisi od preklapanja aktina i miozina, bar ne u onom
rasponu gde se od takvih filimenata ne očekuje da se međusobno preklapaju (dužine veće od
maksimuma krive tetaničkog napona), niti da utiču na unutrašnju brzinu aktiviranja i
normalnog funkcionisanja spojnica. Eksperimenti koji su usledili u kojima je početna dužina
mišića bila izmenjena (bila je veća ili manja od 3,1 μm) nisu izmenili rezultate (Gordon, E.F.
Hoksli i Džujijan, 1966). Ovi zaključci su potrvđeni u eksperimetnima novijeg datuma i
prošireni kako bi pokazali da je brzina skraćivanja neopterećenog mišića na vrhuncu napona
kojji nastaje prilikom grča, u osnovi ista kao ona do koje dolazi usled
tetanusa
(Edman,
1978). Štaviše, ogoljena vlakna koja su delimično aktivirana u rastvoru kalcijuma, verovatno
dostižu maksimalnu brzinu skraćivanja istu kao potpuno aktivirana vlakna u jednakim
uslovima (Tems i saradnici, 1974), iako ova vrednost još uvek nije ustanovljena.
Slika 2. Brzina skraćivanja mišića sa lakim teretom na primeru žabe
Sve su ovo vrlo ubedljivi dokazi u korist ideje da spojnice deluju kao nezavisni
generatori sile. Oni izjednačavaju maksimalnu brzinu skraćivanja sa brzinom skraćivanja
neopterećenog mišića jedne spojnice – više spojnica, paralelno poređanih, mogu povećati
sposobnosti mišića da stvori napon, ali ne mogu preneti brzinu skraćivanja neopterećenog
mišića.
5