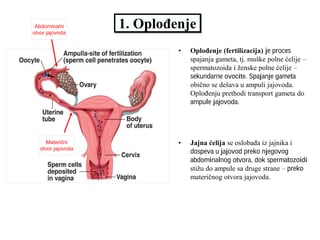
ANDRECEUM I GINECEUM
ANDRECEUM
Skup svih prašnika jednog cveta naziva se andreceum (
androeceum
). Svaki prašnik,
po pravilu, sastoji se od prašničkog konca (filamenta) i prašnice (antera). Antera –
(prašnica) se sastoji iz dva dela – to su poluantere (
thecae
), koje su međusobno razdvojene
sterilnim tkivom, označenim kao konektiv. Svaka teka (poluantera) ima po dve polenove
kesice, u kojima se stvara polen.
Svi delovi cveta su, manje ili više, probraženi listovi. Prema tome i prašnici su
listovi (koji su u znatnoj meri preobraženi). Pošto se u prašnicima obrazuju spore, onda se
takvi listovi nazivaju sporofili. Zbog toga što je kod cvetnica izražena heterosporija
(postoje mikro- i makrospore), tačnije je reći da su prašnici mikrosporofili. Poznato je da
se na mikrosporofilima obrazuju mikrosporangije. U ovom slučaju mikrosporangije su
polenove kesice. U polenovim kesicama (mikrosporangijama) obrazuju se polenova zrna –
mikrospore.
Osnovna funkcija prašnika je obrazovanje polena (cvetnog praha), polenovih zrna.
Ima, međutim, biljaka kod kojih su pojedini prašnici, ili ceo krug prašnika, izgubili
sposobnost da obrazuju polen. Takvi prašnici koji ne obrazuju nikakav polen ili stvaraju
sterilan polen nazivaju se staminodije (sterilni prašnici). Kod nekih ženskih cvetova, koji
vode poreklo od hermafroditnih, nalaze se neplodni prašnici, odnosno staminodije.
Obrazovanje polenovih zrna počinje u polenovim kesicama znatno pre otvaranja
cvetnih pupoljaka. Još u mladoj anteri, u ranim fazama njenog razvića, ćelije sub-
epidermalnog sloja postaju krupnije i dele se tangencijalnim deobama u dva sloja,
unutrašnji će dati arheriosporijum (sporogeno tkivo), a od spoljašnjeg sloja postaće deo
zida polenove kesice.
Ćelije spoljnog sloja dalje se dele i diferenciraju u tri različita sloja, koji
opkoljavaju arhesporijum. Spoljni sloj, koji se nalazi neposredno ispod epidermisa, naziva
se endotecijum (fibrozni sloj). Endotecijum se sastoji iz krupnih ćelija, koje su često mrtve
i sa odrvenelim membranama u definitivnom stanju. Ćelije endotecijuma imaju važnu
ulogu u otvaranju antera. Jedan ili nekoliko slojeva sitnijih ćelija ispod endotecijuma čine
tzv. međusloj. Ovaj sloj se kasnije utroši za ishranu polena. Unutrašnji sloj koji se
neposredno graniči sa arhesporijumom, naziva se tapetum. Tapetum je izgrađen od živih
ćelija, koje su jedno- ili dvojedarne a u nekih biljaka i četvojedarne.
Ćelije arherisporijuma se dele i daju matere ćelije polenovih zrna. Svaka matera
ćelije deli se redukcionom deobom i daje četiri polenova zrna (mikrospore), u stvari četiri
ćelije (tetrade).
Obrazovanje tetrada (mikrospora) vrši se na dva načina. Sukcesivno ili simultano.
Pri sukcesivnom tipu, posle prve mejotične deobe obrazuje se pregradna membrana, koja
deli materu ćeliju na dve ćelije – dijade. Posle druge mejotične deobe ponovo se obrazuje
membrana u svakoj sestrinskoj ćeliji i tako postaju četiri nove ćelije – tetrade mikrospora.
Kod ovog tipa razvića mikrospora između prve i druge mejotičke deobe nastupa interfaza
koja, u zavisnosti od uslova, može biti i prilično duga. U slučaju simultanog tipa razvića
mikrospora ne obrazuje se membrana posle prve mejotičke deobe, već odmah nastupa
druga, tj. jedro se deli na dva pa na četiri, a zatim se od periferije ka centru protoplasta
matere ćelije javljaju ubori, kojima se ona podeli na četiri nove ćelije.
Sukcesivni tip razvoja tetrada mikrospora je češće posmatran u monokotiledonih
biljaka, a simultani u dikotila (sreće se i u nekih monokotila:
Juncaceae,
Cyperaceae,
Orchidaceae
). Predpostavlja se da je sukcesivni tip progresivniji, a simultani primitivniji.
Od tipa obrazovanja zvisi kako će mikrospore biti raspoređene u tetradama.
1
Proces formiranja mikrospora iz matere ćelije naziva se mikrosporogeneza.
Posle rastvaranja membrane matere ćelije, oko mikrospora počinje da se obrazuje
sopstvena membrana i mikrospora se preobraća u polenovo zrno. U početku polenovo zrno
ima gustu citoplazmu, bez jasno izražene vakuolizacije, a jedro je smešteno u sredinu
ćelije. Sa povećanjem prometa materija u citoplazmi polenovog zrna postepeno se formira
vakuola, koja potiskuje jedro polenovog zrna prema ćelijskom zidu.
Oko polenovog zida obrazuju se dve membrane-spoljna egzina i unutrašnja intina.
Egzina je uglavnom celulozna, a ponekad je i kutinizovana. Na spoljnoj strani egzine često
se nalaze različite skulpture, koje su karakteristične za familije, rodove i vrste. Stoga u
egzini se nalaze pore, preko kojih se obezbeđuje veza polenovih zrna sa okolnom
sredinom. Broj pora je različit i zavisi od osobine egzine. Ako je egzina glatka onda je broj
pora mali (1-3), a ako je ona debela i sa skulpturama broj pora dostiže i 30 do 40
Unutrašnja opna-intina je veoma tanka, dvoslojna je i sastavljena obično od
celuloze i pektinskih materija.
Posle formiranja zida polenovog zrna počinje deoba jedra mikrospore, koja uvek
protiče na nekoliko dana pre otvaranja cveta. Kao rezultat ove deobe nastaju dve, po
veličini različite ćelije. Jedna ćelija je veća, s krupnim okruglim jedrom u kome je krupno
jedarce i sa žitkom vakuoliziranom citoplazmom. To je vegetativna ćelija. Ona odgovara
jako reduciranom mikroprotalijumu, odnosno ona je gametofit. Druga manja ćelija, s
visokim sadržajem RNK, naziva se generativna ćelija (anteridijalna ćelija). U početku je
genrativna ćelija priljubljena uz jedan deo zida polenovog zrna, a kasnije u toku njenog
razvića ona prodire dublje u unutrašnjost polenovog zrna, zadobija vretenastu formu i
smešta se u citoplazmu vegetativne ćelije. Ovakvim položajem obezbeđeno je bolje
rastenje i razviće generativne ćelije na račun vegetativne. Generativna ćelija ima veoma
tanku opnu u vidu dvojne membrane. Ona se satoji od pektinskih materija. U citoplazmi se
nalaze obično plastidi (leukoplasti, ređe hloroplasti), mitohondrije i ribozomi. Citoplazma
vegetativne ćelije je bogata rezervnim materijama, od kojih su najviše rasprostranjeni
lipidi, polisaharidi (skrob), aminokiseline, nukleinske kisline (RNK), belančevinasti
kristali, vitamini.
Generativna ćelija se još u samom polenovom zrnu može podeliti na dve ćelije-
spermije ili spermatične ćelije (to su muški gameti), ili se ova deoba vrši tek pri klijanju
polenovog zrna, u polenovoj cevi. Prema tome, obrazovanje muških gameta vrši se ili u
polenovom zrnu, ili u polenovoj cevi. Proces obrazovanja muških gametofita naziva se
gametogeneza. Muški gameti, spermije, sadrže više ili manje plazme koja okružuje jedro.
Forma i veličina spermije jako varira u različitih vrsta (okrugle, ovalne, sočivaste,
elipsoidne, vretenaste i dr.). Forma spermija se menja u toku njihove ontogeneze. Mlade
spermije imaju više okruglu, a zrele-izdužene formu. Spermije mogu biti sitne, srednje i
krupne. Za dikotile su tipične sitne (ređe, srednje), a za monokotile krupne i srednje.
Zrela polenova zrna sadrže ugljene hidrate (šećer, skrob), lipide, belančevine,
aminokiseline, nukleinske kiseline, fermente (citohromoksidazu, peroksidazu, i dr.),
vitamine, karotinoide, mineralne soli. Primena histohemijskih metoda pri izučavanju zrelih
polenovih zrna u 70 vrsta angiospermi (iz 41 familije) pokazala je relativnu promenljivost
u sadržaju rezervnih i fizioloških aktivnih materija i to ne samo u okviru jedne vrste već i
iste antere.
Produkcija polena po jednoj biljci je veoma velika, naročito u anemofilnih biljaka.
Tako, na primer, jedna biljka kukuruza u proseku proizvodi oko 50.000.000 polenovih
zrna.
Veličina polena varira u širokim granicama, u zavisnosti od vrste biljaka, od 0,008
mm (u
Ficus elaticas
) do vrlo krupnih, kao npr. u tikve – do 0,2 mm. Boja polenovih zrna
2

da se sinteza belančevina i nukleinskih kiselina vrši intenzivno u gornjem delu mikrospore
(megaspore). Zbog polarnih razlika u prometu materije u makrospori, kasnije se javlja
različit stepen životnih sposobnosti jedra koja se nalaze bliže mikropili, odnosno bliže
halazi. Sledeća etapa razvića embrionove kesice predstavlja u stvari nastanak
makroproprotalijuma (makrogametofita). To se vrši tako što se makrospora koja se često
naziva i “jednojedarna embrionova kesa” dalje razvija na taj način što se njeno jedro deli
na dva koja odlaze ka suprotnim polovima, jedno ka mikropili, a drugo ka halazi. Zatim se
svako od ovih jedara deli da bi se na kraju dobilo po četiri jedra na svakom polu. Znači,
dobija se jedan osmojedarni stadijum u formiranju embrionove kesice. Uporedo sa
deobama jedara vrši se i povećanja veličine embrionove kesice, kao i delimično
vakuoliziranje njene citoplazme. Dalji proces razvitka embrionove kesice odnosno
makrogametofita vrši se nat način što po jedno jedro sa svakog pola odlazi u centralni deo
embrionove kese. Na polovima ostaju po tri jedra i svako od njih sa odgovarajućom
citoplazmom daje posebnu ćeliju. Na taj način na mikropilarnom polu nastaje jajni aparat,
koji se sastoji od jedne krupnije – jajne ćelije i dve sitnije sinergide, dok se na halaznom
polu formiraju tri ćelije – antipode. Ona dva jedra koja su otišla ka centru stapaju se ili
samo priljubljuju jedno uz drugo. Na taj način postaje takozvano sekundarno (centralno)
jedro embrionove kesice, koje se najvećim delom citoplazme gradi centralnu ćeliju.
Opisanim procesima ne samo da se formirao od megaspore makrogametofit već se
formirao i makrogamet (jajna ćelija), odnosno izvršena je i makrogametogeneza.
U mikropilarnom delu, gde se nalazi jajni aparat, vrši se najintenzivnija sinteza
belančevina i maksimalna koncentracija RNK. U halaznom delu embrionove kesice, u
rejonu antipoda, često se vrše procesi redukcije.
Sinergide su po svojoj građi i veličini ćelije istog tipa. One su kruškaste ili izdužene
forme. Jedro se u sinergidama nalazi u gornjem delu gde je skoncentrisana citoplazma,
dok se u njihovom donjom delu obrazuje vakuola. Sinergide imaju bitnu ulogu u
privlačenju polenove cevi prema embrionovoj kesici i jajnom aparatu, a takođe doprinose
rastvaranju mambana polenove cevi, pošto sadrže razne fermente, kao citazu, pektinazu.
Jajna ćelija obično zauzima središni položaj u jajnom aparatu. Ona je krupnija od
sinergida, izdužene ili kruškaste forme, ima krupno jedro u svom donjem delu i gustu
citoplazmu. U gornjem delu jajne ćelije formira se vakuola. U citoplazmi jajne ćelije
nalaze se ribozomi, mitohondrije i plastidi (leukoplasti). Ona je bogata polisaharidima i
lipidima. U njoj se nagomilavaju u većoj količini RNK i belančevine, pri čemu se najveća
količina RNK nalazi u donjem delu jajne ćelije i usredsređuje se oko jedra, a najmanje je
nje u gornjem delu jajne ćelije. Dakle, jajna ćelija ima jasno izraženu polarnost. U
halaznom delu embionove kesice, kao što je već pomenuto, obrazuju se tri ćelje, antipode.
Embrionova kesica koja se razvija iz jedne makrospore (megaspore), dok ostale tri
ćelije nastale mejotičkom deobom, degenerišu, spada u grupu monospornih embrionovih
kesica. Monosporne emrionove kesice mogu biti dvojakog tipa u zavisnosti od toga da li
vrše tri, ili samo dve uzastopne mitotičke deobe njihovih jedara. U prvom slučaju nastaje
osmojedarni stadijum embrionove kesice, tako da se u zreloj embrionovoj kesici, kao što je
izneto, obrazuje šest haploidnih ćelija (jajna ćelija, dve sinergide i tri antipode) i jedna
sekundarna ili centralna (diploidna) ćelija čije jedro nastaje sjedinjavanjem dvaju polarnih
jedara (sekundarno jedro). To je
Polygonum
– tip embrionove kesice koji je široko
rasprostranjen u angiospermi, pa se zbog toga naziva i normalni tip.
Pored grupe monospornih, postoje i grupe dispornih i tetraspornih embrionovih
kesica u čijem razviću učestvuju dve odnosno četiri makrospore (megaspore).
4
Oplođenje
Kada polenovo zrno padne na žig tučka, počinje proces oplođenja. Kod nekih biljkaka
oplođenje nastupa već 30' od oprašivanja, a kod nekih drugih ovaj proces traje i nekoliko
dana, nedelja ili godina. Kada polenovo zrno padne na žig tučka ono počinje da klija.
Unutrašnja opna polenovog zrna, intina, izdužuje se u polenovu cev i izrasta kroz pore na
egzini. Polenova cev raste od žiga do semenog zametka. Ona se izdužuje i prodire kroz
stubić, ili kroz specijalizovano »transportno tkivo«. Sa izduživanjem i prodiranjem
polenove cevi, sa njom se kreće i sadržaj polenovog zrna (vegetativna i generativna ćelija).
Generativna ćelijase ovde deli na dve spermatične ćelije. Kada dođe do plodnika, polenova
cev raste kroz njegovo tkivo i najkraćim putem se upravlja ka otvoru na semenom
zametku, tj. Ka mikropili. Takav pravac rastenja, kada polena cev prodire u semeni
zametak kroz mikropilu naziva se porogamija. Ukoliko polenova cev prodire kroz halazu
to se naziva halazogamija. Polenova cev raste vrhom, i sa tim porastom sadržaj cevi se
stalno pomera ka vrhu, dok se stari delovi cevi guraju od vrha polenove cevi i pregrađuju
kaloznim pregradama. Neke od „najbržih“ polenovih cevi u
in vivo
uslovima je od
1,8mm/sec kod noćurka do 2,7 mm/sec kod kukuruza, dok je kod nekih ovaj porast dosta
sporiji. Kada polenova cev stigne u dodir sa opnom embrionove kesice, ona se rastvara.
Tog momenta izlazi sadržaj polenove cevi, ali ne dospeva direktno na jajnu ćeliju, već
pored nje ili u jednu od sinergida, koajse usled toga razori. Vegetativno jedro degeneriše,
jedna spermatična ćelija se sjedinjuje sa jajnom ćelijom a druga spermatična ćelija sa
centralnom ćelijom. Na ovaj način obavljeno je dvojno oplođenje. Oplođenjem jajne ćelije
nastaje diploidni zigot, a iz oplođene sekundarne ćelije nastaje triplodni endosperm.
Analiza citogenetičkih uzroka steriliteta
Citogenetički sterilitet je izražen u jabuka, krušaka, višanja, vinove loze, šljive i drugim
voćnim vrstama i sortama vinove loze.
Radi utvrđivanja citoloških uzroka muške sterilnosti koristimo arhisporijalne ćelije
muškog gametofita. Arhisporijalne ćelije muškog gametofita nalaze se u mladim anterama
cveta.
Ukoliko je proces redukcione deobe normalan, onda u tom slučaju ne treba i očekivati
sterilnost formiranog muškog gametofita. Najčešće nenormalnoti za vreme redukcione
deobe su:
1.
formiranje univalenata ili konjugacija polivalentnih hromozoma u
PROFAZI I, Mejoze I
2.
zaostajanje hromozoma na deobnom vretenu u ANAFAZI I, mejoze I, i
3.
formiranje umesto normalnih tetrada, diade, triade, pentade, heksade i dr.
u TELOFAZI II.
4.
CITOMIKSIS
Redukciono deljenje arhisporijalnih ćelija se dešava u različitim voćnim vrstama u
različitim vremenskim periodima. Uglavnom koštičave voćne vrste počinju ranije
redukcionu deobu. Redukciona deoba u ovim voćnim vrstama se zapaža i prema spoljnim
manifestacijama, jer se obično uvećavaju cvetni pupoljci. Međutim, u krušaka redukciona
deoba se kasnije i poznaje se po boji antera. Ukoliko je zelena boja u potpunosti izražena
onda je znak da redukciona deoba još nije počela. Međutim, prelaskom zelene u ružičastu
boju znak je da je redukciona deoba u toku, ukoliko su antere crvene boje onda je znak da
je redukciona deoba završena. U jabuka se može utvrditi vreme početka redukcione deobe ,
takođe po spoljnim manifestacijama. Redukciono deljenje je prisutno u cvetovima jabuka
kada se pojave prvi zeleni listići na cvetnim pupoljcima.
5

znatna razlika u veličini polena, s tim što je u diploida polen obično ujednačen po obliku i
veličini, a u triploida ima i sitnih polenovih zrna. Heterogenost oblika i veličine polena
jednog genotipova ukazuje na anomalije u mikrosporogenezi, nejednaku podelu
hromozoma i obrazovanje poliada. Iste anomalije se uočavaju i kod genotipova nastalih
hromozomnim mutacijama.U mutanata jednostrukih recipročnih translokacija izražen je
semisterilitet i lako se poznaju analizom polena koji je oko 50% normalne klijavosti
ishodne forme, a po veličini i obliku 50% je normalan, a 50% sitna i smežurana polenova
zrna.
Analiza oblika i veličine polena može se obaviti bojenjem acetokarminom, metodom u
visećoj kapi saharoze, ali je najpogodnija u imerzionom ulju /kedrovom ulju/, pri kome je
najmanje bubrenje polena, a time i adekvatniji rezultati.
Analiza steriliteta usled nenormalnosti ženskog gametofita
Postoje više tipova ženskog gametofita: monosporni, disporni i tetrasporni, a ovi imaju
podtipove koji se razlikuju po načinu formiranja embrionove kesice.
U voćaka i vinovoj lozi obrazuju se tzv. normalni tip, monosporni, koji je zastupljen u
70% ispitivanih drugih angiospermi. Ovaj tip embrionove kesice obrazuje se posle druge
redukcione deobe formiranja makrospore, koja se posle deli sa 3 uzastopne deobe i daje
normalnu osmojedarnu embrionovu kesicu.
Kao i pri obrazovanju polena tako i pri makrosporogenezi mogu u raznih sorata voćaka
i vinove loze doći do izražaja citološki uzroci steriliteta. Oni se javljaju u vidu raznih
anomalija pri redukcionoj deobi, a analogne su anomalijama koje se odigravaju pri
mikrosporogenezi, pa se zbog lakšeg postupka i većeg broja ćelija pri mikrisporogenezi, pa
se zbog lakšeg postupka i većeg broja ćelija muškog arhesporija vrši samo analiza
nenormalnosti pri redukcionoj deobi materinskih ćelija polena, a indirektno se zaključuje o
nenormalnosti pri obrazovanju makrospora. Međutim, pri obrazovanju ženskog gametofita
posle formiranja makrospore mogu doći do izražaja anomalije kao posledica neregularne
redukcione kao i druge dodatne anomalije. Za analizu razvoja ženskog gametofita i
anomalija koje mogu dovesti do steriliteta najpodesnija je parafinska metoda spravljanja
mikroskopskih preparata sa debljinom preseka /uzdužnog preseka semenog začetka na
mikrotomu/ od 15 mikrona i konbinovanim bojenjem, šafraninom, hematoksilinom,
gencijalna violet i oranže ge uz diferenciranje boje u karanfilinskom ulju.
Od velikog broja cvetova jabuka i kruška zametne plod samo 5-10%. Analizom
embrionove kesice može se videti da prema položaju cveta u cvasti i tipu rodne grančice
jabuke ima različiti udeo smetnji i anomalija u njihovom razvoju. Ove anomalije se mogu
svesti u više grupa: u vreme punog cvetanja nije formirana osmojedarna embrionova
kesica, atrofirana izvesna jedra ili ćelije embrionove kesice- antipode sinergide, jajna ćelija
ili sekundarna ćelija embrionove kesice.
Pored toga u vreme punog cvetanja mogu se naći dve ili više embrionove kesice u
istom semenom zametku, tako da međusobno otežavaju dalji razvoj. pri obrazovanju
većeg broja embrionovih kesica u istom semenom začetku, obično su u raličitim fazama
razvitka, a u vreme otvaranja kruničnih listića izvestne mogu biti još nerazvijene i
funkcionalno nesposobne, a druge pak da je zahvatila atrofija. Atrofija embrionove kesice,
delimična ili potpuna može se konstatovati, ako je do nje došlo na mikroskopskim
preparatima, kao i nenormalnosti u ranijem razvoju materinske ćelije makrospore i njenom
redukcionom deljenju. Na bazi frekfencije pomenutih anomalija može se izračunati stepen
smanjenja fertiliteta ispitivane sorte. Smetnje u razvoju ženskog gametofita su veće
frefencije u triploidnih nego u diploidnih sorata jabuka i krušaka.
7